07 CATALYTIC MECHANISMS
07 CATALYTIC MECHANISMS
Executive Summary
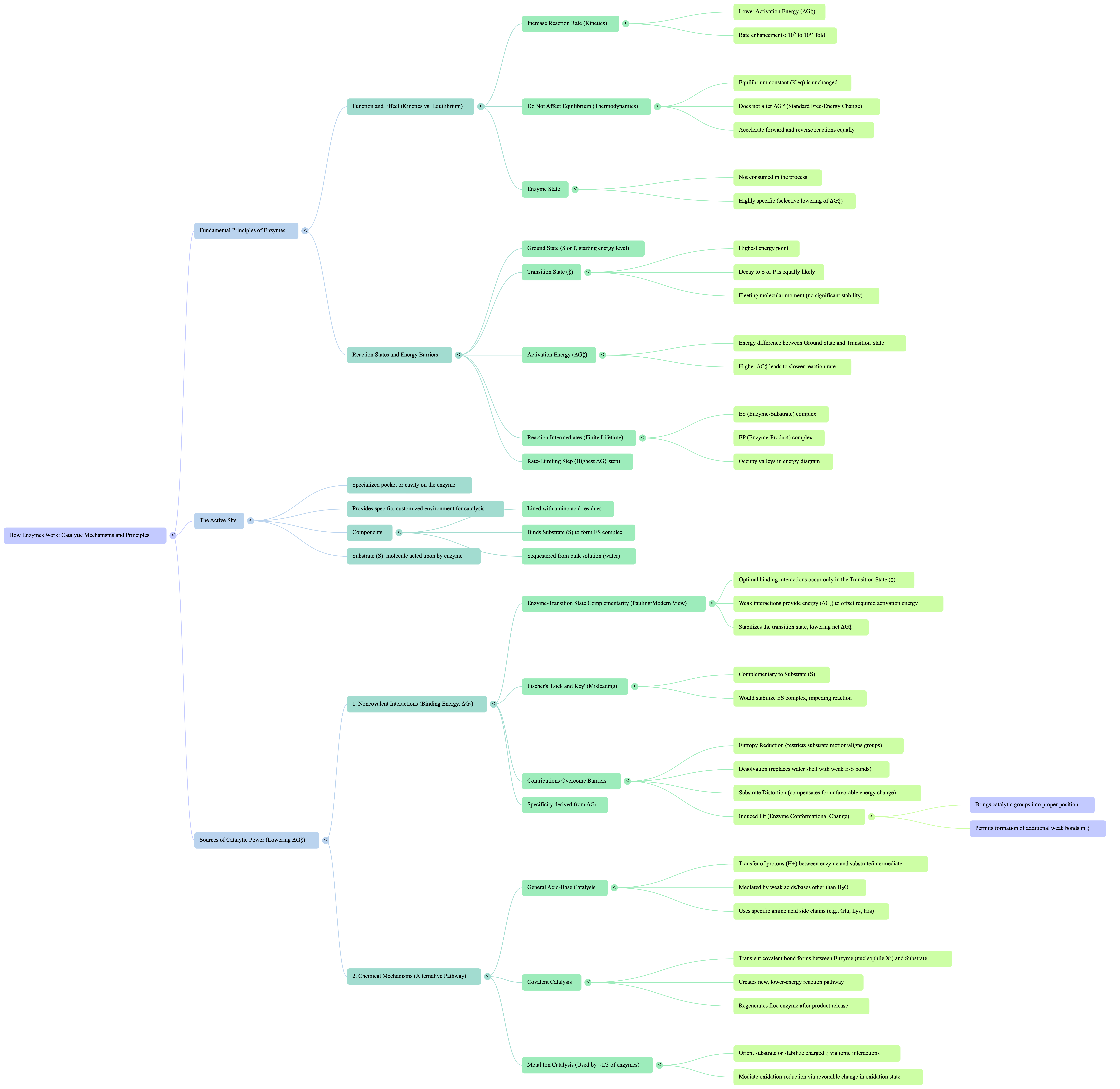
Enzymes are the essential biological catalysts that make life possible. While the molecules within cells are generally stable, the chemical reactions required for processes like digestion, energy production, and nerve signaling would occur far too slowly on their own to sustain an organism. Enzymes solve this problem by dramatically accelerating reaction rates by 5 to 17 orders of magnitude. They achieve this by lowering the activation energy (ΔG‡)—the energy barrier that must be overcome for a reaction to proceed. It is a fundamental principle of catalysis that while enzymes increase reaction rates, they do not alter the final reaction equilibrium. An enzyme helps a reaction reach its natural balance point much faster but does not change what that balance point is.
The primary source of an enzyme's catalytic power is binding energy (ΔGB). This is the free energy released when the enzyme forms multiple weak, noncovalent interactions (such as hydrogen bonds and ionic interactions) with the substrate. Crucially, enzymes have evolved not to bind the starting substrate molecule as tightly as possible, but rather to be structurally complementary to the high-energy transition state of the reaction. This means the strongest binding interactions are formed only when the substrate is distorted into this fleeting, unstable state. The favorable energy released from these interactions effectively "pays for" the energy required to reach the transition state, resulting in a much lower net activation energy and an extraordinary increase in reaction speed. To complement this powerful strategy, enzymes also employ specific chemical mechanisms, including general acid-base catalysis, covalent catalysis, and metal ion catalysis, to further facilitate the chemical transformation of the substrate.
--------------------------------------------------------------------------------
1. The Fundamental Role of Enzymes in Biological Systems
1.1. Introduction to Enzymatic Catalysis
In the aqueous, neutral-pH, and mild-temperature environment inside a living cell, most biological molecules are quite stable. While this stability is essential for maintaining cellular structures, it poses a challenge for metabolism. The chemical reactions necessary for life—from digesting food to sending nerve signals—would proceed at rates far too slow to be useful without assistance. Enzymes are nature's solution to this problem. They are highly efficient and specific biological catalysts that accelerate chemical reactions, often by 5 to 17 orders of magnitude, to rates compatible with the speed of life.
1.2. The Active Site: The Hub of Catalytic Activity
An enzyme-catalyzed reaction takes place within a specialized pocket or cleft on the enzyme's surface. The key components of this process are:
- Enzyme: The biological catalyst, typically a large protein, which participates in the reaction but is not consumed by it.
- Active Site: The specific region on the enzyme where the substrate binds and the chemical reaction occurs. As shown in the structure of the enzyme chymotrypsin (Figure 6-1), the active site provides a unique microenvironment, with key amino acid residues precisely positioned to facilitate catalysis.
- Substrate (S): The molecule that binds to the active site and is chemically transformed by the enzyme.
- Product (P): The molecule resulting from the enzymatic transformation of the substrate.
- Enzyme-Substrate (ES) and Enzyme-Product (EP) Complexes: These are transient, intermediate states where the substrate or product is bound to the enzyme. Unlike the fleeting transition state, these complexes represent real, albeit short-lived, molecular species that occupy energy minima (valleys) on the reaction pathway, as shown in Figure 6-3. They represent distinct steps along the pathway, as formalized in the general equation for an enzymatic reaction (Equation 6-1): E + S ⇌ ES ⇌ EP ⇌ E + P
1.3. The Core Principle: Affecting Rates, Not Equilibria
The primary function of any catalyst, including an enzyme, is to increase the rate of a reaction. It is equally important to understand what catalysts do not do: they do not affect reaction equilibria. A reaction is at equilibrium when the concentrations of reactants and products are no longer changing. An enzyme achieves this state much more rapidly by accelerating both the forward (S → P) and the reverse (P → S) reactions to the same degree. The final ratio of products to substrates at equilibrium remains exactly the same as it would be in the absence of the enzyme.
This fundamental distinction between rates and equilibria provides the framework for understanding how enzymes achieve their catalytic power, which is rooted in the principles of thermodynamics and kinetics.
--------------------------------------------------------------------------------
2. The Language of Reactions: Kinetics and Thermodynamics
2.1. Understanding Reaction Progress
To grasp how enzymes function, it is essential to visualize the energy changes that occur as a reactant is converted into a product. Reaction coordinate diagrams are invaluable tools for this purpose. These diagrams plot the free energy of the system against the progress of the reaction, allowing us to clearly distinguish between the factors that determine the reaction's overall energy balance and those that govern its speed.
2.2. Key Thermodynamic Concepts
By referencing a typical reaction coordinate diagram (Figures 6-2 and 6-3), we can define the essential terms that describe the energetic landscape of a reaction:
- Ground State: This refers to the starting free energy level of the system, representing the average energy of the substrate (S) or the product (P) under a given set of conditions.
- Transition State (‡): This is not a stable chemical species but a fleeting molecular moment at the very peak of the energy barrier. At this point, bond breakage, bond formation, and charge development have progressed to the precise state from which the molecule is equally likely to decay back to the substrate or proceed forward to the product.
- Activation Energy (ΔG‡): This is the energy barrier for the reaction. It is defined as the difference in free energy between the ground state of the substrate and the high-energy transition state. A fundamental principle of kinetics is that a higher activation energy corresponds to a slower reaction rate.
- Standard Free-Energy Change (ΔG'°): This value represents the difference in the ground-state free energies of the product and the substrate, measured under standard biochemical conditions (pH 7.0). A negative ΔG'° indicates that the reaction is exergonic, meaning the products are more stable than the reactants and the reaction is thermodynamically favorable.
2.3. Linking Thermodynamics to Reaction Outcomes
These thermodynamic concepts are directly linked to the observable characteristics of a chemical reaction.
- The standard free-energy change (ΔG'°) determines the reaction equilibrium (K'eq). As shown in Table 6-4, there is a direct mathematical relationship between these values. A large, negative ΔG'° signifies that the equilibrium constant (K'eq) will be much greater than 1, meaning the reaction at equilibrium will strongly favor the formation of products.
- The activation energy (ΔG‡) determines the reaction rate constant (k). According to transition-state theory (Equation 6-6), the relationship between these two is both inverse and exponential. This is the most critical takeaway for understanding catalysis: even a small decrease in the activation energy leads to a dramatic increase in the reaction rate.
These principles explain what needs to happen for a reaction to speed up; next, we explore how enzymes specifically manipulate these energy barriers.
--------------------------------------------------------------------------------
3. The Power of Interaction: Binding Energy and Transition-State Theory
3.1. The Source of Catalytic Energy
The extraordinary catalytic power of enzymes comes from a single primary source: binding energy (ΔG_B). This is the free energy released upon the formation of numerous weak, noncovalent interactions—such as hydrogen bonds, ionic interactions, and the hydrophobic effect—between the enzyme and its substrate. To put this in context, a decrease in activation energy of just 5.7 kJ/mol results in a 10-fold increase in reaction rate. The energy from a single weak interaction is typically 4 to 30 kJ/mol, and the sum of multiple such interactions can easily provide the 60 to 100 kJ/mol reduction in activation energy needed to explain the massive rate enhancements observed for many enzymes. This binding energy is the fundamental currency that the enzyme uses to lower the activation energy of the reaction.
3.2. Beyond "Lock and Key": The Transition-State Complementarity Model
- In 1894, Emil Fischer proposed the "lock and key" hypothesis, suggesting that an enzyme's active site was a rigid structure perfectly complementary to its substrate. While this idea was influential, it is incorrect for explaining catalysis.
- An enzyme that is perfectly complementary to its substrate would be a very poor catalyst. As illustrated by the "stickase" analogy in Figure 6-5b, such an enzyme would bind and stabilize the starting substrate molecule in its ground state. This stabilization would create a deep energy trough, making it more difficult for the substrate to reach the high-energy transition state and thus increasing the activation energy and slowing the reaction.
- The modern and correct model posits that enzymes are complementary to the reaction's transition state. The active site is structured to bind the transition-state version of the substrate with the highest possible affinity. While some weak interactions form when the substrate first binds, the full complement of stabilizing interactions is formed only when the substrate is distorted into the high-energy transition state.
3.3. How Binding Energy Lowers Activation Energy
The core mechanism of enzyme catalysis lies in a simple energetic trade-off. The reaction itself has an inherently unfavorable energy barrier, the positive activation energy (+ΔG‡), that must be overcome. The enzyme facilitates this by providing favorable binding energy, a negative free-energy change (–ΔG_B), that is maximized in the transition state.
As shown in Figure 6-6, the binding energy released by the optimal enzyme-substrate interactions in the transition state "pays for" a significant portion of the energy required to reach that state. The summation of the unfavorable activation energy and the favorable binding energy results in a much lower net activation energy for the enzyme-catalyzed reaction. This dramatic reduction in the energy barrier is the direct cause of the massive rate enhancements observed in enzymatic reactions.
This principle is powerfully illustrated by the enzyme triose phosphate isomerase. In the reaction it catalyzes, the chemical rearrangement occurs at carbons 1 and 2 of the substrate. However, interactions between the enzyme and the substrate's phosphate group at carbon 3—which is not directly involved in the chemical change—account for over 80% of the catalytic rate acceleration. This shows that the entire binding interaction, not just at the point of bond breaking, contributes to lowering the activation energy.
3.4. The Unity of Catalysis and Specificity
The principle of transition-state stabilization not only explains an enzyme's catalytic power but also its remarkable specificity. Catalysis and specificity arise from the same phenomenon. An enzyme active site is a highly precise three-dimensional structure with functional groups arranged optimally to form a rich network of weak interactions with a particular substrate in its transition state. This same exquisitely tuned site will inherently be unable to form such a strong network of interactions with the transition state of a different, competing molecule. Specificity is therefore a direct consequence of an active site optimized for the transition state of one particular reaction.
--------------------------------------------------------------------------------
4. Harnessing Binding Energy: Specific Catalytic Strategies
4.1. Overcoming Reaction Barriers
The abstract concept of using binding energy to lower the activation barrier translates into overcoming several distinct physical and thermodynamic hurdles that slow down reactions in solution. These include: (1) the entropy of molecules in solution, (2) the solvation shell of water surrounding the substrate, (3) the energy required for substrate distortion, and (4) the need for proper alignment of catalytic groups. Binding energy is strategically used to conquer each of these barriers.
4.2. Mechanisms Driven by Binding Energy
- Entropy Reduction: In solution, molecules move randomly, and the probability of two reactants colliding with the perfect orientation for a reaction is very low. By binding substrates within the confines of the active site, an enzyme severely restricts their freedom of motion and locks them into the precise orientation required for reaction. This entropy reduction dramatically increases the effective concentration of the reactants and the likelihood of a productive collision. Studies of model reactions show this effect alone can be responsible for rate enhancements of 10⁵ to 10⁸-fold.
- Desolvation: Most biomolecules in the cell are surrounded by a shell of ordered water molecules (a solvation shell) that can interfere with a reaction. The formation of weak bonds between the enzyme and substrate is energetically favorable enough to displace this water shell. This desolvation process strips away the water molecules, removing a potential impediment to the reaction.
- Substrate Distortion: For many reactions to occur, the substrate must be forced into a strained or distorted conformation that mimics the transition state. This distortion requires an input of energy. The binding energy released from enzyme-substrate interactions in the transition state compensates for this energy requirement, helping to push the substrate toward its reactive form.
- Induced Fit: Contrary to the rigid "lock and key" model, both the enzyme and substrate are flexible. The initial binding of a substrate often induces a conformational change in the enzyme, a phenomenon known as induced fit. This structural change can reposition critical amino acid side chains in the active site, bringing catalytic groups into the perfect alignment to act on the substrate. This adjustment also frequently allows for the formation of additional, stronger binding interactions that further stabilize the transition state.
These physical strategies are often complemented by mechanisms where the enzyme takes an even more direct chemical role in the reaction.
--------------------------------------------------------------------------------
5. A Deeper Look: Chemical Catalytic Mechanisms
5.1. Actively Participating in the Reaction
In addition to harnessing binding energy, many enzymes use mechanisms in which functional groups within the active site—such as amino acid side chains or bound metal ions—actively and directly participate in the chemical transformation. These strategies often involve the formation of transient covalent bonds with the substrate or the transfer of protons to and from the substrate to stabilize fleeting intermediates.
5.2. Common Chemical Strategies
- General Acid-Base Catalysis:
- This mechanism involves the transfer of a proton either to or from the substrate or a reaction intermediate. Its primary role is to stabilize unstable charged intermediates that form during a reaction, preventing them from collapsing back to reactants.
- It is crucial to distinguish between specific acid-base catalysis, which uses only the H+ or OH- ions from water, and general acid-base catalysis, which uses weak acids and bases (such as the ionizable side chains of amino acids) as proton donors or acceptors.
- This strategy is incredibly common and is employed by the vast majority of enzymes.
- Covalent Catalysis:
- In this mechanism, a reactive functional group on the enzyme forms a transient covalent bond with the substrate.
- This process creates an alternative, lower-energy reaction pathway involving an enzyme-substrate intermediate. For this strategy to be effective, both the formation and the subsequent breakdown of this covalent intermediate must be faster than the uncatalyzed reaction.
- Metal Ion Catalysis:
- Metal ions play several critical roles in catalysis. They can: (1) help orient the substrate correctly for the reaction through ionic interactions; (2) stabilize charged transition states electrostatically; and (3) mediate oxidation-reduction reactions by undergoing reversible changes in their own oxidation state.
- The importance of this mechanism is highlighted by the fact that nearly a third of all known enzymes require one or more metal ions to be catalytically active.
In practice, most enzymes are sophisticated molecular machines that combine the overarching principle of transition-state stabilization via binding energy with one or more of these specific chemical strategies to achieve their remarkable catalytic efficiency.
Resources
- MIND_MAP
- FLASHCARDS
- SELF-ASSESSMENT
- PRESENTATION_SLIDES
{kind=link}