08 MICHAELIS-MENTEN KINETICS
08 MICHAELIS-MENTEN KINETICS
Executive Summary
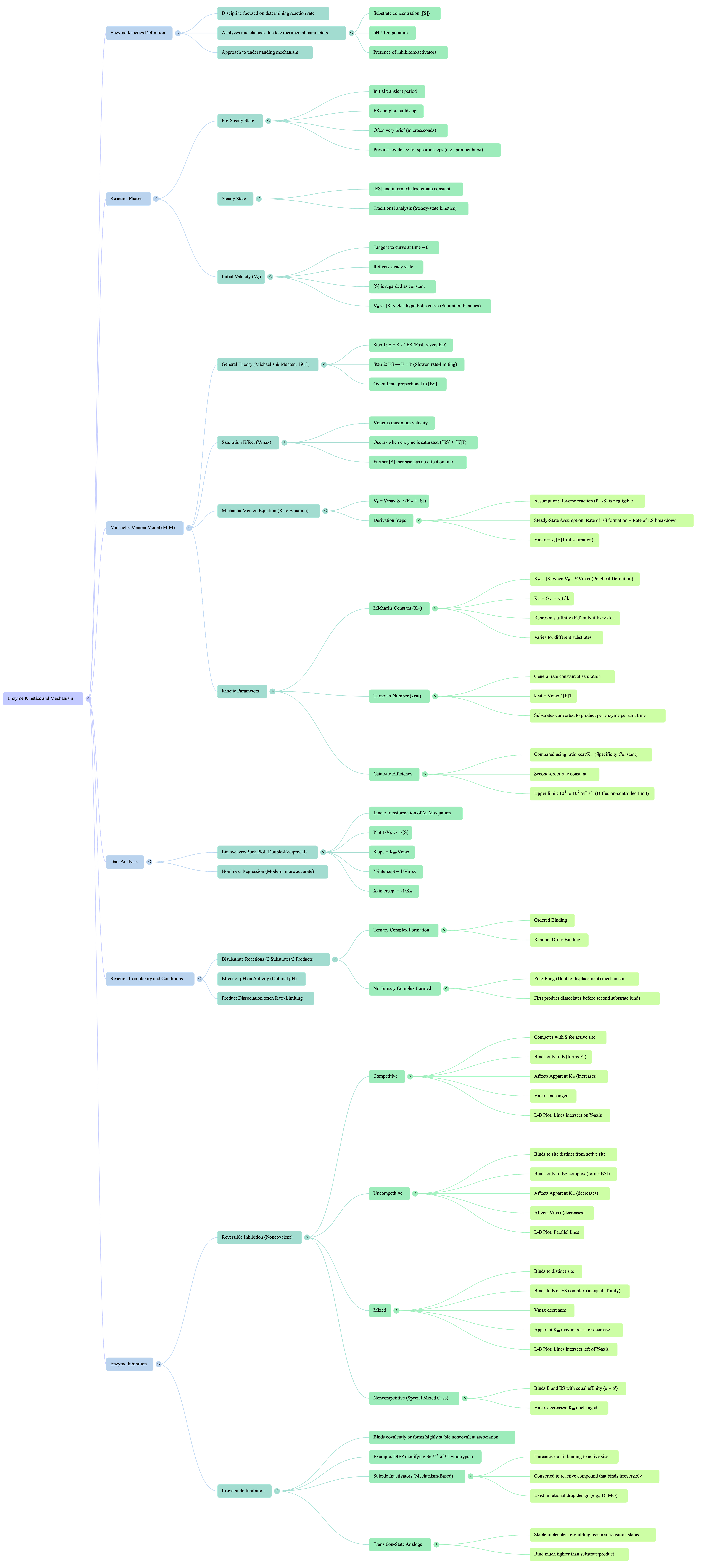
Enzyme kinetics is the quantitative study of the rates of enzyme-catalyzed reactions. This field provides critical insights into how enzymes function, complementing knowledge gained from structural and genetic analyses. At its core, the study of kinetics seeks to understand the relationship between the concentration of a substrate—the molecule an enzyme acts upon—and the speed, or velocity, of the reaction.
A key observation is that this relationship is not linear. As substrate concentration increases, the reaction velocity initially rises rapidly but eventually levels off, approaching a maximum velocity (Vmax). This plateau occurs because the enzyme becomes saturated; that is, all available enzyme molecules are bound to substrate and are working at their maximum capacity. Adding more substrate at this point cannot make the reaction go any faster.
The Michaelis-Menten model, proposed in 1913, provides the foundational mathematical description for this behavior. It is built on the concept of a two-step process: the rapid and reversible formation of an enzyme-substrate (ES) complex, followed by a slower, rate-limiting step where the complex breaks down to form the product and regenerate the free enzyme.
This relationship is quantified by the Michaelis-Menten equation, which uses two key parameters:
- Vmax (Maximum Velocity): The theoretical maximum rate of the reaction when the enzyme is fully saturated with substrate.
- Km (The Michaelis Constant): The substrate concentration at which the reaction proceeds at half of its maximum velocity (½Vmax). It is an indicator of an enzyme's efficiency at low substrate concentrations.
Furthermore, enzyme activity can be modulated by inhibitors—molecules that slow or stop catalysis. These are vital in medicine, as many drugs function as enzyme inhibitors. They are classified by how they interact with the enzyme and substrate, with distinct types like competitive, uncompetitive, and mixed inhibitors altering Vmax and Km in characteristic ways.
Ultimately, the Michaelis-Menten model remains a cornerstone of biochemistry. It provides an essential framework for characterizing enzyme performance, comparing catalytic efficiencies, and understanding the mechanisms of metabolic regulation and drug action.
--------------------------------------------------------------------------------
1. The Foundations of Enzyme Kinetics
While modern techniques like structural biology provide a static snapshot of an enzyme's form, enzyme kinetics offers a dynamic view of its function. It is the discipline focused on determining the rate of a reaction and how it changes in response to changes in experimental parameters. This approach remains a primary and essential method for elucidating an enzyme's mechanism of action, providing a quantitative description of its catalytic power and efficiency that complements static structural data.
To analyze these reactions, biochemists focus on several key concepts that simplify the process:
- Enzyme Kinetics: The study of reaction rates and how they change in response to factors like substrate concentration, pH, and the presence of inhibitors.
- Pre-Steady State: This is the initial, very brief period (often lasting only microseconds) immediately after an enzyme and substrate are mixed. During this phase, the concentration of the enzyme-substrate (ES) complex rapidly builds up.
- Steady State: Following the pre-steady state, the reaction enters a much longer phase where the concentration of the ES complex remains relatively constant. This occurs because the rate at which the ES complex is formed is balanced by the rate at which it breaks down to form product. The traditional analysis of reaction rates, known as steady-state kinetics, focuses on this period.
- Initial Velocity (V₀): This is the reaction rate measured at the very beginning of the reaction (time = 0), typically when less than 2-3% of the substrate has been converted to product. Measuring the initial velocity offers a critical advantage: at this point, the change in substrate concentration [S] is negligible, allowing it to be treated as a constant. This simplifies the analysis greatly, even though this initial rate measurement reflects steady-state conditions.
By systematically measuring the initial velocity (V₀) under steady-state conditions across a range of different initial substrate concentrations, biochemists can construct a kinetic profile that quantitatively characterizes an enzyme's performance. This profile reveals the fundamental relationship between the amount of available substrate and the rate at which an enzyme can function.
2. The Michaelis-Menten Model: Substrate Concentration and Reaction Rate
The relationship between substrate concentration [S] and the initial reaction velocity (V₀) is a defining characteristic of enzyme catalysis. Unlike uncatalyzed reactions where the rate is directly proportional to reactant concentration, enzyme-catalyzed reactions exhibit a hyperbolic dependence. The foundational theory explaining this phenomenon, proposed by Leonor Michaelis and Maud Menten in 1913, is centered on the formation of an enzyme-substrate (ES) complex as a necessary step in catalysis.
As the concentration of substrate [S] increases from very low levels, the initial velocity (V₀) shows a distinct pattern of behavior that leads to enzyme saturation:
- At low [S], the initial velocity increases in a nearly linear fashion with the addition of more substrate.
- At higher [S], the rate begins to level off, with each increase in substrate concentration producing a smaller and smaller increase in velocity.
- Eventually, the velocity approaches a plateau, a theoretical maximum rate that cannot be surpassed, no matter how much more substrate is added.
This saturation effect is explained by the finite number of enzyme molecules available. At the plateau, the enzyme is working at its full potential. The key terms associated with this phenomenon are:
- Maximum Velocity (Vmax): This is the theoretical maximum rate the reaction can achieve. Vmax is reached when the enzyme is "saturated," a state where virtually every enzyme molecule is bound to a substrate molecule, forming the ES complex.
- Enzyme-Substrate (ES) Complex: The formation of this noncovalent complex between the enzyme and its substrate is the central postulate of the model. Its concentration determines the overall reaction rate.
The Michaelis-Menten model simplifies the catalytic process into a two-step reaction:
- Fast, Reversible Binding: The enzyme (E) and substrate (S) rapidly and reversibly combine to form the enzyme-substrate complex (ES). E + S ⇌ ES
- Slower, Rate-Limiting Catalysis: The ES complex breaks down in a slower step to release the product (P) and regenerate the free enzyme (E). ES → E + P
Because the second step is the slower of the two, it acts as a bottleneck and is considered the rate-limiting step of the overall reaction. It is important to note that the full second step is also reversible (ES ⇌ E + P). However, for the derivation of the Michaelis-Menten equation, the reverse reaction of product forming substrate is ignored. This is a valid simplifying assumption because at the very beginning of the reaction (t=0), the product concentration is negligible.
3. Quantifying Enzyme Performance: The Michaelis-Menten Equation
The characteristic hyperbolic curve observed when plotting initial velocity (V₀) against substrate concentration [S] can be described with precision by an algebraic formula: the Michaelis-Menten equation. This equation is the cornerstone of steady-state kinetics, as it provides a direct, quantitative relationship between the reaction rate and the concentrations of substrate and enzyme.
The Michaelis-Menten equation is expressed as:
V₀ = (Vmax * [S]) / (Km + [S])
Each component of this equation has a specific, measurable meaning:
- V₀: The initial velocity of the reaction at a given substrate concentration.
- Vmax: The maximum velocity the reaction can achieve when the enzyme is completely saturated with substrate.
- [S]: The initial concentration of the substrate.
- Km (The Michaelis Constant): This constant has a crucial operational definition: Km is the substrate concentration at which the reaction velocity is exactly half of Vmax (V₀ = ½Vmax). Km is a composite of multiple rate constants ((k₋₁ + k₂)/k₁) that describe the formation and breakdown of the ES complex. While related to substrate affinity, it is not a direct measure of it. Only in the specific scenario where product formation is much slower than substrate dissociation (k₂ << k₋₁) does Km approximate the dissociation constant (Kd), a true measure of affinity.
The Michaelis-Menten equation is derived using the steady-state assumption. This assumption posits that during the steady-state phase of the reaction, the rate at which the ES complex is formed is equal to the rate at which it is broken down (either by dissociating back to E and S or by proceeding forward to E and P). This allows the concentration of the ES complex to be expressed in terms of measurable quantities, leading to the final equation.
The Lineweaver-Burk Plot
To more easily determine the values of Vmax and Km from experimental data, the Michaelis-Menten equation can be algebraically transformed into a linear form. The most common transformation is the Lineweaver-Burk equation:
1/V₀ = (Km/Vmax)(1/[S]) + 1/Vmax
This equation has the form of a straight line (y = mx + b). By plotting the reciprocal of velocity (1/V₀) versus the reciprocal of substrate concentration (1/[S]), known as a double-reciprocal plot, researchers can obtain a straight line. The kinetic parameters can then be determined directly from the intercepts of this line:
- The y-intercept is equal to 1/Vmax.
- The x-intercept is equal to -1/Km.
While the Lineweaver-Burk plot is valuable for visualizing kinetic data and diagnosing inhibition mechanisms, modern nonlinear regression methods are now more common and offer accuracy superior to Lineweaver-Burk. This is because the double-reciprocal transformation tends to give undue weight to data obtained at low substrate concentrations and can distort experimental errors. These fundamental parameters, Km and Vmax, provide the basis for more advanced metrics used to compare the overall efficiency of different enzymes.
4. Advanced Metrics of Catalytic Efficiency
While Km and Vmax are essential for characterizing the behavior of a single enzyme, comparing the overall catalytic efficiency of different enzymes, or the same enzyme with different substrates, requires more universal parameters. The "gold standard" metrics for this purpose are the turnover number (kcat) and the specificity constant (kcat/Km).
- kcat (The Turnover Number): This parameter represents the maximum number of substrate molecules that a single enzyme molecule can convert into product per unit of time, assuming the enzyme is fully saturated with substrate. It is a direct measure of an enzyme's maximal catalytic speed and is calculated from Vmax and the total enzyme concentration, [E]T: kcat = Vmax / [E]T.
- kcat/Km (The Specificity Constant): This ratio is considered the most effective measure for comparing the catalytic efficiency of enzymes. It represents the apparent second-order rate constant for the reaction when the substrate concentration is much lower than Km ([S] << Km), a condition often found in the cell. The specificity constant reflects an enzyme's ability to both bind its substrate (related to Km) and convert it to product (related to kcat), thereby providing a comprehensive measure of its overall performance.
Catalytic Perfection
The rate at which an enzyme can operate has an upper physical limit determined by the rate of diffusion, which is the fastest that an enzyme and its substrate can encounter each other by random motion in an aqueous solution. This diffusion-controlled limit is between 10⁸ and 10⁹ M⁻¹s⁻¹. Enzymes with a kcat/Km value in this range are considered to have achieved catalytic perfection. They have evolved to catalyze their reaction so efficiently that the rate is limited only by how quickly they can find their substrate. Examples of such enzymes include Catalase, Fumarase, and Acetylcholinesterase, all of which operate near this theoretical maximum speed.
5. Modulation of Enzyme Activity: Inhibition
Enzyme inhibitors are molecules that interfere with catalysis, slowing or even completely halting enzymatic reactions. They are of immense importance in biology and medicine, forming the basis for a vast number of pharmaceutical drugs. Beyond their therapeutic applications, the study of enzyme inhibitors has also provided valuable information about enzyme mechanisms and has helped define metabolic pathways. Enzymologists use the characteristic kinetic effects of different inhibitors as diagnostic tools to deduce mechanistic details. Inhibitors are broadly divided into two main classes: reversible and irreversible.
Reversible Inhibition
Reversible inhibitors can be distinguished by their effect on the enzyme's kinetic parameters, Vmax and Km. There are three primary types:
- Competitive Inhibition
- Mechanism: The inhibitor is often structurally similar to the substrate and competes directly with it for binding to the enzyme's active site. The binding of the inhibitor and the substrate is mutually exclusive.
- Effect on Kinetics: A competitive inhibitor increases the apparent Km of the enzyme, meaning a higher concentration of substrate is required to achieve ½Vmax. However, it does not change Vmax. At sufficiently high substrate concentrations, the substrate can outcompete the inhibitor, allowing the enzyme to reach its normal maximum velocity.
- Lineweaver-Burk Plot: The lines for inhibited and uninhibited reactions intersect at the y-axis (at 1/Vmax).
- Clinical Example: A classic example of competitive inhibition is the treatment for methanol poisoning. Methanol is converted by the enzyme alcohol dehydrogenase to toxic formaldehyde. To prevent this, a high concentration of ethanol is administered intravenously. Ethanol competes with methanol for the active site of alcohol dehydrogenase, slowing formaldehyde production and allowing the kidneys to safely excrete the methanol.
- Uncompetitive Inhibition
- Mechanism: The inhibitor binds at a site distinct from the active site, and it binds only to the enzyme-substrate (ES) complex. It does not bind to the free enzyme.
- Effect on Kinetics: An uncompetitive inhibitor decreases both the apparent Vmax and the apparent Km by the same factor. Vmax decreases because some enzyme is effectively removed from the reaction as the inactive ESI complex. Km decreases because the inhibitor stabilizes the ES complex, reducing the substrate concentration needed to reach half-maximal velocity.
- Lineweaver-Burk Plot: The lines are parallel.
- Mixed Inhibition
- Mechanism: The inhibitor binds to a site distinct from the active site, but it can bind to either the free enzyme (E) or the ES complex.
- Effect on Kinetics: A mixed inhibitor always decreases the apparent Vmax. Its effect on the apparent Km varies; it can either increase or decrease depending on whether the inhibitor has a higher affinity for the free enzyme or the ES complex. A special case of mixed inhibition, known as noncompetitive inhibition, occurs when the inhibitor binds to E and ES with equal affinity. In this specific case, Vmax is decreased, but the apparent Km remains unchanged.
- Lineweaver-Burk Plot: The lines intersect to the left of the y-axis, but not on either axis.
Irreversible Inhibition
Irreversible inhibitors permanently inactivate an enzyme by forming a stable covalent bond with an essential functional group in the active site or by forming a very tight, non-dissociating noncovalent association.
A highly specific and potent class of these are suicide inactivators (or mechanism-based inactivators). These molecules are designed to be unreactive until they are acted upon by their target enzyme. The enzyme's own catalytic mechanism transforms the inactivator into a highly reactive compound that then covalently attaches to and permanently inactivates the enzyme.
- Clinical Example: The drug difluoromethylornithine (DFMO) is a suicide inactivator used to treat African sleeping sickness, a disease caused by trypanosome parasites. DFMO is a "biochemical Trojan Horse" that targets the parasite's ornithine decarboxylase enzyme, which is essential for cell division but more stable than its human counterpart. The enzyme processes DFMO as if it were the normal substrate, but in doing so, converts DFMO into a reactive species that covalently bonds to and inactivates the enzyme. This highly specific action kills the parasite with minimal effect on the human host.
6. Summary: Key Concepts for Retention
This section distills the most critical, high-yield concepts from the study of Michaelis-Menten kinetics, designed for effective studying and long-term retention.
- Saturation Kinetics The velocity of an enzyme-catalyzed reaction does not increase infinitely as more substrate is added. Instead, it approaches a maximum rate (Vmax) as the enzyme becomes saturated with substrate. This hyperbolic relationship between substrate concentration and reaction velocity is a hallmark of enzyme catalysis.
- The Michaelis-Menten Equation This behavior is mathematically described by the core equation of enzyme kinetics: V₀ = (Vmax[S])/(Km + [S]). This formula quantitatively links initial velocity (V₀), maximum velocity (Vmax), and substrate concentration ([S]) through the Michaelis constant (Km).
- The Meaning of Km and Vmax These two parameters provide a practical description of an enzyme's performance. Vmax is the enzyme's maximum speed when it is working at full capacity (saturated). Km is the substrate concentration required for the enzyme to work at half its maximum speed (½Vmax). A lower Km generally indicates that the enzyme can operate efficiently even at low substrate concentrations.
- Comparing Enzyme Efficiency To compare the catalytic efficiency of different enzymes or of one enzyme with different substrates, the kcat/Km ratio (specificity constant) is the most informative metric. It accounts for both the rate of catalytic turnover (kcat) and substrate binding (related to Km), providing a measure of overall performance.
- Competitive vs. Other Forms of Reversible Inhibition Understanding how inhibitors work is vital for pharmacology and biochemistry. Competitive inhibitors bind to the active site, competing with the substrate; they increase the apparent Km but do not affect Vmax. Uncompetitive and Mixed inhibitors bind to sites other than the active site and always decrease Vmax. Uncompetitive inhibitors decrease both Vmax and Km, while Mixed inhibitors always decrease Vmax but can either increase or decrease Km. This kinetic distinction is fundamental to identifying and designing effective drugs.
Resources
- MIND_MAP
- FLASHCARDS
- SELF-ASSESSMENT
- PRESENTATION_SLIDES
{kind=link}