12 GENES & CHROMOSOMES
12 GENES AND CHROMOSOMES - A LEARNING SUMMARY
Executive Summary
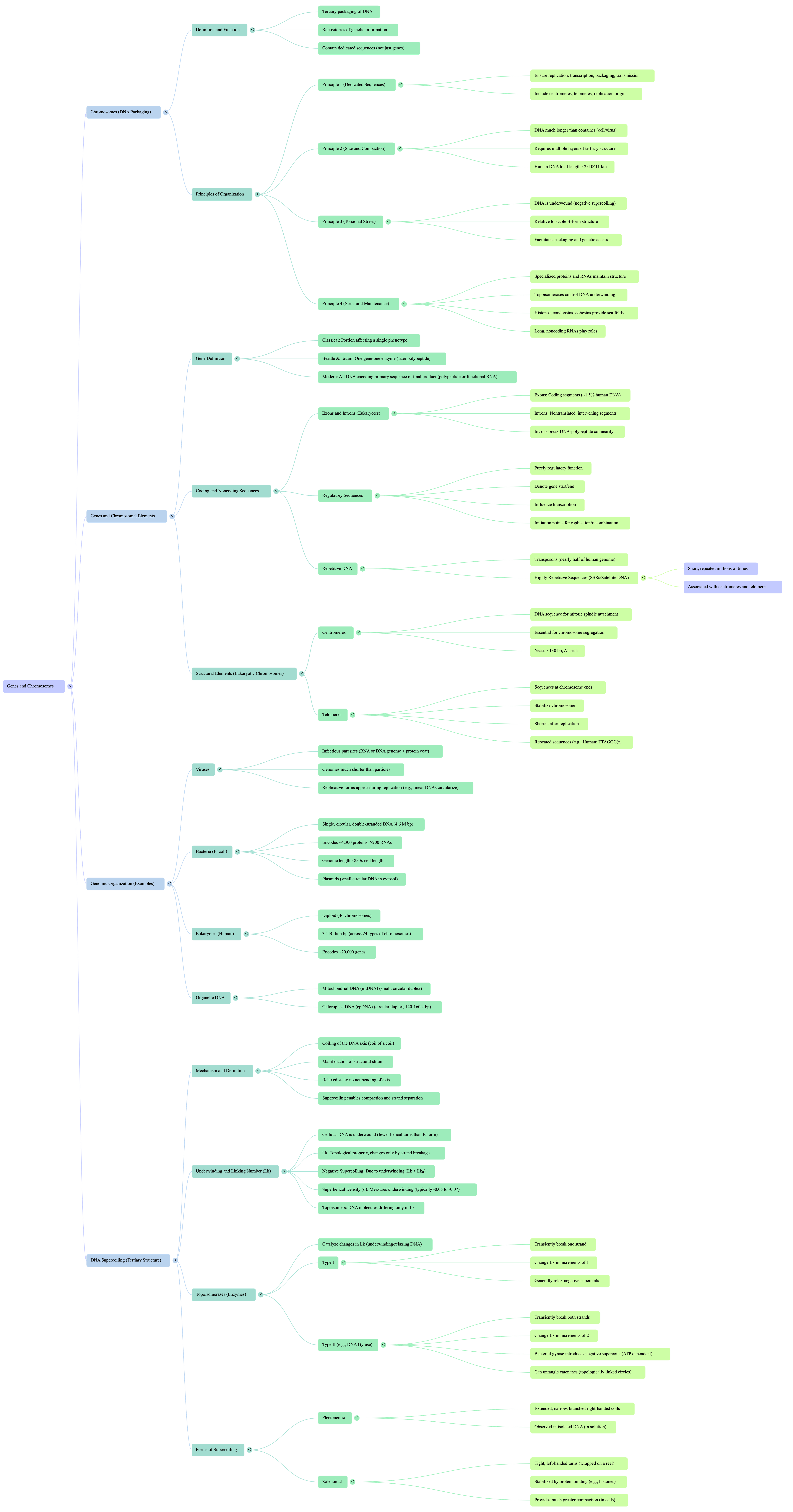
Every living organism faces a fundamental challenge of information management: how to package an immense length of DNA into a microscopic cellular space while keeping the genetic blueprint accessible for use. This summary explores the elegant, multi-layered solution to this problem, centered on the structure and organization of genes and chromosomes.
The modern biochemical definition of a gene is the entire stretch of DNA that codes for a final product, which can be either a protein or a functional RNA molecule. This genetic information is stored on DNA molecules that are orders of magnitude longer than the cells or viruses containing them. For instance, the DNA of a single human cell, if stretched out, would be two meters long. To solve this packaging problem, life has evolved sophisticated chromosomal structures. In prokaryotes like E. coli, the genome typically consists of a single, circular chromosome. In contrast, eukaryotes have much larger genomes divided among multiple linear chromosomes, which possess unique structural features. These include non-coding introns that interrupt the coding exons, a central centromere essential for cell division, and protective telomeres at the chromosome ends.
A key physical principle governing DNA compaction in all organisms is supercoiling. Cellular DNA is typically kept in an underwound state, meaning it has fewer helical turns than its most stable relaxed form. This creates torsional strain that causes the DNA to twist upon itself, forming supercoils. This "negative supercoiling" not only compacts the DNA but also stores energy that makes it easier for the two strands to separate—a critical step for DNA replication and transcription. In essence, chromosome structure is not a static storage unit but a dynamic, highly regulated system that masterfully balances the need for extreme compaction with the necessity of accessing genetic information to sustain life.
--------------------------------------------------------------------------------
1. The Fundamental Unit of Heredity: Defining the Gene
To understand the complex architecture of chromosomes, we must first examine their fundamental functional unit: the gene. Our scientific conception of a gene has evolved significantly over the last century. What began as an abstract unit of heredity, linked to observable traits, is now understood as a precise molecular entity with a defined biochemical function and structure.
1.2 The Evolving Definition of a Gene
The journey to our modern understanding of the gene reflects major breakthroughs in genetics and biochemistry.
- Classical Definition: Initially, a gene was defined by its effect on a visible property, or phenotype, such as the gene that determines eye color. This was a functional definition based on observation, long before the molecular nature of heredity was known.
- One Gene-One Enzyme Hypothesis: A pivotal shift occurred with the work of George Beadle and Edward Tatum in the 1940s. By studying mutant strains of fungi, they demonstrated that a specific gene codes for a single enzyme. This concept was later refined to the "one gene-one polypeptide" hypothesis, acknowledging that many proteins consist of multiple polypeptide chains, each encoded by a separate gene.
- Modern Biochemical Definition: Today, a gene is defined with biochemical precision as all the DNA that encodes the primary sequence of a final gene product. This product can be a polypeptide chain or a functional RNA molecule with a structural or catalytic role.
1.3 Beyond the Code: Regulatory Sequences
In addition to the coding regions, chromosomes contain vast stretches of DNA known as regulatory sequences. These segments do not code for a final product but are essential for controlling and orchestrating the processes of life.
- Marking Boundaries: They signal the beginning and end of genes, ensuring that cellular machinery reads the correct segments of DNA.
- Controlling Transcription: They act as switches, influencing when and to what extent a gene is transcribed into RNA.
- Initiating Key Processes: They serve as specific starting points for DNA replication and recombination.
While we now understand the gene as a discrete unit of information, the true biological challenge lies in organizing and managing millions of these units encoded in a DNA molecule thousands of times longer than the cell itself—a problem of physical data compression.
2. The Packaging Problem: Comparing Genomes Across Life
Every organism, from the simplest virus to the most complex eukaryote, must solve a universal biological puzzle: how to fit an enormous length of DNA into a microscopic container. The strategies employed to meet this challenge vary across the domains of life, reflecting differences in genome size, complexity, and cellular architecture.
2.2 The Scale of the Challenge
The discrepancy between the length of a DNA molecule and the size of the compartment that holds it is staggering. Chromosomal DNA is consistently orders of magnitude longer than the cell or virus it resides in.
- The circular chromosome of the bacterium E. coli has a contour length of about 1.7 mm, which is approximately 850 times the length of the cell itself.
- The DNA from a single human diploid cell, containing 46 chromosomes, would measure about 2 meters if all the molecules were laid end-to-end.
2.3 Genome Organization in Viruses and Bacteria
- Viruses: As infectious parasites, viruses package their genetic material (either RNA or DNA) into a small protein coat. The contour length of their DNA is typically hundreds of times longer than the viral particle. During replication inside a host, many viral genomes adopt different structures called replicative forms; for example, a linear DNA molecule may become circular to facilitate its replication.
- Bacteria: The genome of a typical bacterium like E. coli consists of a single, circular, double-stranded DNA molecule. In addition to this main chromosome, many bacteria also contain plasmids—small, independent circular DNA molecules. Plasmids often carry useful genes, such as those conferring resistance to antibiotics, and can be transferred between bacteria.
2.4 Genome Organization in Eukaryotes
Eukaryotic genomes are generally much larger and more complex than those of prokaryotes. This genetic material is organized into multiple, linear chromosomes housed within a nucleus.
- Size and Gene Count: The E. coli genome contains approximately 4.6 million base pairs (bp) and encodes about 4,300 proteins and over 200 functional RNAs. In contrast, the human genome has 3.1 billion bp distributed across 24 different chromosome types but encodes only approximately 20,500 proteins.
- Complexity vs. Size: This comparison highlights a key finding in genomics: genome size does not directly correlate with an organism's complexity or its number of genes. This surprising lack of correlation is partly explained by the vastly different internal structure of eukaryotic genes, which contain extensive non-coding regions.
- Organellar DNA: Eukaryotic cells also contain small, circular DNA molecules within their mitochondria (mtDNA) and, in plants, their chloroplasts (cpDNA). This is a legacy of their evolutionary origin as free-living bacteria.
The greater size and division of genetic labor in eukaryotes necessitate a more intricate chromosomal architecture compared to their prokaryotic counterparts.
3. The Intricate Architecture of Eukaryotic Chromosomes
The complexity of eukaryotic life is mirrored in the sophisticated structure of its chromosomes. Beyond simply being long strands of DNA, eukaryotic chromosomes contain specialized elements that are essential for accurate gene expression, replication, and the faithful transmission of genetic material during cell division.
3.2 Interrupted Genes: Exons and Introns
A defining feature of most eukaryotic genes is their segmented nature. But what does this mean biochemically? It means the DNA sequence that codes for a product is not continuous but is interrupted by non-coding regions.
- Exons are the segments of a gene that contain the coding information for the final product.
- Introns are the non-coding, intervening DNA segments.
This intron-exon structure is common in eukaryotes but rare in bacteria. Introns can be extensive; for example, in the gene for ovalbumin (an egg protein), the seven introns make up 85% of the gene's total DNA. In humans, it is estimated that only about 1.5% of our total DNA consists of protein-coding exons. The immediate consequence of this structure is that the initial RNA copy of the gene must be precisely edited—or 'spliced'—to remove the introns before the message can be translated into a protein.
3.3 Essential Structural Elements
For a linear chromosome to be stable and function correctly, it requires several key structural components.
- Centromeres The centromere is a specific DNA sequence that functions as an attachment point for proteins that link the chromosome to the mitotic spindle during cell division. Its biological importance is paramount: this connection ensures the orderly and equal segregation of chromosomes into the two daughter cells, preventing genetic abnormalities. These regions are often rich in highly repetitive DNA sequences known as simple sequence repeats, which contribute to their unique structural identity.
- Telomeres Telomeres are specialized structures found at the physical ends of linear eukaryotic chromosomes. They consist of thousands of tandem repeats of a short, specific DNA sequence, generally in the form of (TxGy)n on one strand and (AxCy)n on the other. In humans, this sequence is (TTAGGG)n. Like centromeres, telomeres are composed of simple sequence repeats. Their primary function is to stabilize the chromosome, protecting the ends from degradation and preventing them from being recognized by the cell as broken DNA.
These essential structural elements—introns, centromeres, and telomeres—define a eukaryotic chromosome's identity and function. However, for this intricate architecture to fit within the nucleus, it must be subjected to extreme physical compression, a process governed by the physics of DNA supercoiling.
4. The Physics of Compaction: DNA Supercoiling
To fit within a cell, the immense length of a DNA molecule must be coiled upon itself in a process called supercoiling. This is not a random tangle but a highly regulated process that introduces torsional strain into the DNA double helix. This strain is fundamental to both compacting the genome and regulating access to the genetic information it contains.
4.2 Underwinding and Torsional Strain
The foundational concept of supercoiling is that most DNA inside a cell is underwound. This means it has fewer helical turns than it would in its most stable, relaxed B-form.
This underwinding creates structural strain. To relieve this strain, the axis of the DNA double helix twists on itself, forming a supercoil. This state, known as negative supercoiling, is energetically favorable for the cell because the stored torsional energy makes it easier to separate the two DNA strands. Strand separation is a prerequisite for essential processes like DNA replication and the transcription of genes into RNA.
4.3 Quantifying Supercoiling: The Linking Number (Lk)
The degree of supercoiling is described by a topological property called the Linking Number (Lk). Think of Lk as a fixed property of a closed loop of rope made of two intertwined strands. You cannot change the number of times the strands cross over each other without cutting one of them. In the same way, the linking number of a closed-circular DNA molecule (or a linear one constrained by proteins) cannot change unless an enzyme, a topoisomerase, temporarily breaks a strand.
- In simple terms, Lk is the number of times one DNA strand winds around the other in a closed-circular molecule.
- The process of underwinding decreases the linking number relative to that of relaxed DNA (a state where Lk < Lk₀). This negative change in Lk directly results in the formation of negative supercoils.
4.4 Enzymatic Regulation: Topoisomerases
Cells precisely control the level of supercoiling using a class of enzymes called topoisomerases. These enzymes regulate DNA topology by changing the linking number.
- Type I Topoisomerases: These enzymes work by creating a transient break in one DNA strand, allowing the other strand to pass through before resealing the break. This changes the Lk in increments of 1.
- Type II Topoisomerases: These enzymes create a transient break in both DNA strands, passing another segment of double-stranded DNA through the gap before resealing. This changes the Lk in increments of 2. The bacterial enzyme DNA gyrase is a notable Type II topoisomerase that actively introduces negative supercoils into DNA, a process that requires the energy of ATP.
4.5 The Two Forms of Supercoils
Imagine two ways to store a long extension cord. You could twist it back on itself into a series of branching loops—this is plectonemic supercoiling. It's stable but still occupies a lot of space. Alternatively, you could wrap it tightly and orderly around a reel—this is solenoidal supercoiling. This second method achieves far greater compaction and is how the cell, using protein 'reels,' organizes its DNA.
- Plectonemic: This is an extended, right-handed supercoil that forms in isolated, protein-free DNA in a solution.
- Solenoidal: This is a tight, left-handed coil where the DNA is wrapped around a central axis. While less stable on its own, this solenoidal form is stabilized within the cell by binding to architectural proteins. In eukaryotes, the primary example is the wrapping of DNA around histone proteins to form nucleosomes, which achieves a much greater degree of compaction than the plectonemic form.
Ultimately, the genome is not a static library but a dynamic molecular machine, where the interplay between defined chromosomal architecture, specialized regulatory proteins, and the stored energy of supercoiling achieves the remarkable feat of securely packaging life's blueprint while ensuring it remains readily accessible for expression and replication.
Resources
- MIND_MAP
- FLASHCARDS
- SELF-ASSESSMENT
- PRESENTATION_SLIDES
{kind=link}