13 STRUCTURES of CHROMOSOMES
13 STRUCTURE OF CHROMOSOMES
Executive Summary
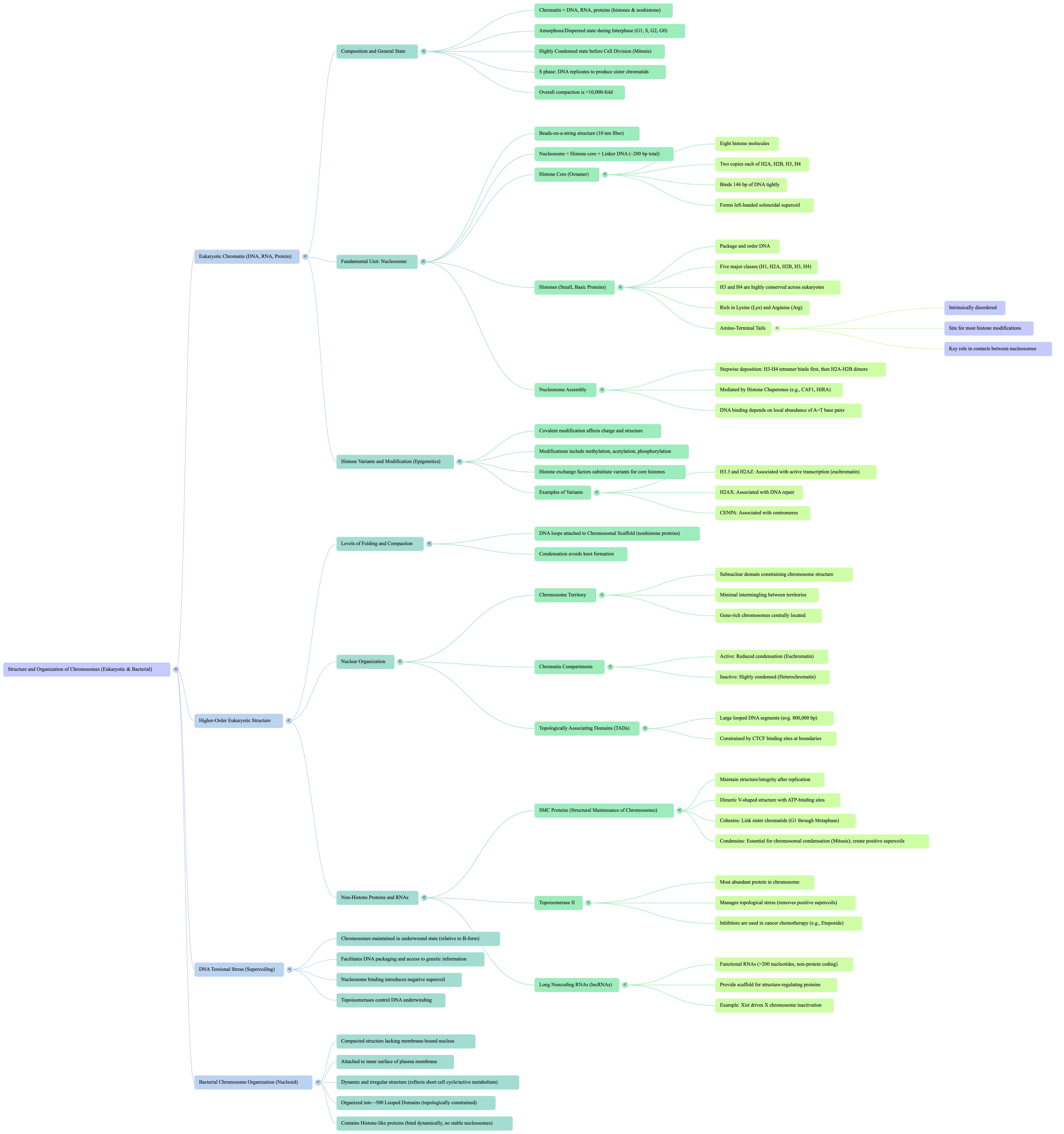
The packaging of genetic material presents a remarkable biological challenge: fitting several meters of DNA into a cell nucleus that is mere micrometers in diameter. This feat, equivalent to packing kilometers of thread into a small ball, requires a sophisticated and hierarchical system of organization. The fundamental material of eukaryotic chromosomes is chromatin, a complex of DNA, structural proteins, and RNA. The primary architects of this structure are a class of small, positively charged proteins called histones.
The first and most fundamental level of DNA compaction is the nucleosome, in which a segment of DNA wraps around a core of eight histone proteins, creating a structure often described as "beads-on-a-string." This initial step compacts the DNA approximately sevenfold. These nucleosomes are then folded into progressively more complex structures. Higher-order organization involves arranging the DNA into large loops, which are further segregated within the nucleus into functionally distinct compartments, such as transcriptionally active euchromatin and inactive, condensed heterochromatin. Ultimately, this multi-layered folding achieves a compaction of over 10,000-fold, culminating in the highly condensed chromosomes visible during cell division (mitosis).
Crucially, chromatin is not a static structure. Its organization is highly dynamic, constantly changing to accommodate the cell's needs. The structure decondenses to allow for DNA replication and gene expression during the cell's growth phase (interphase) and condenses dramatically for segregation during mitosis. This dynamic regulation, orchestrated by specialized proteins and RNA molecules, is essential for protecting the genome, ensuring its faithful inheritance, and controlling the flow of genetic information.
--------------------------------------------------------------------------------
1. The Fundamental Challenge: Packaging Genetic Material
The sheer scale of the genetic blueprint within a eukaryotic cell poses an immense physical problem. A typical human cell nucleus, only 5 to 10 micrometers in diameter, must contain DNA molecules that would stretch for meters if laid end to end. To manage this extraordinary length, the cell employs a highly organized and multi-layered packaging system. This system not only makes it possible for the DNA to fit within the nucleus but is also essential for protecting the DNA from damage, regulating gene activity, and ensuring the accurate segregation of genetic material during cell division.
- 1.1. Defining the Core Concepts: Chromosome and Chromatin
- Chromosome: This term has a dual meaning. It can refer to the nucleic acid molecule itself, which serves as the repository of an organism's genetic information. It also refers to the distinct, densely colored bodies that become visible with a light microscope in the nucleus of a eukaryotic cell as it prepares to divide.
- Chromatin: This is the substance that makes up eukaryotic chromosomes. It is a complex of fibers containing DNA, proteins (in approximately equal mass), and a significant amount of RNA. During the non-dividing phases of the cell cycle, chromatin appears as an amorphous, dispersed material within the nucleus.
This fundamental material, chromatin, is built from specific molecular components that direct its intricate folding.
2. The Building Blocks of Chromatin: Histones and Nucleosomes
The elegant organization of DNA within the nucleus is not an intrinsic property of the DNA molecule itself but is orchestrated by a suite of specialized proteins. Chromatin is a highly structured DNA-protein complex, and the principal proteins responsible for the first level of organization are the histones. These proteins act as molecular spools around which the long thread of DNA is wound, forming the fundamental repeating unit of chromatin.
- 2.1. Analysis of Histones: The Master DNA Spools Histones are a family of proteins with specific biochemical properties that make them perfectly suited for their role in packaging DNA.
- Biochemical Nature: Histones are relatively small proteins, with molecular weights ranging from 11,000 to 21,000 daltons. They are exceptionally rich in the basic amino acids arginine and lysine, which are positively charged. This high positive charge allows histones to bind tightly to the negatively charged phosphate backbone of the DNA molecule through strong electrostatic interactions.
- Major Classes: There are five major classes of histones found in all eukaryotic cells: H1, H2A, H2B, H3, and H4.
- Conservation: The amino acid sequences of histones H3 and H4 are remarkably conserved across all eukaryotes, suggesting that their function is so fundamental that it has tolerated very little change over evolutionary time. For example, the H4 histone in peas and cows differs by only two out of 102 amino acids. In contrast, histones H1, H2A, and H2B show more sequence variation between species.
- Modifications: Histones are subject to a wide array of enzymatic modifications, including methylation, acetylation, ADP-ribosylation, phosphorylation, glycosylation, SUMOylation, or ubiquitination. These modifications alter the charge and shape of the histones, which in turn affects the structure of the chromatin and plays a critical role in regulating processes like transcription.
- 2.2. The Nucleosome: The First Level of Compaction The nucleosome is the fundamental organizational unit of chromatin. When partially unfolded, chromatin reveals a "beads-on-a-string" structure, where each "bead" is a nucleosome core particle and the "string" is the DNA connecting them.
- The Histone Core: The core of each nucleosome is an octamer containing eight histone molecules: two copies each of H2A, H2B, H3, and H4.
- The Wrapped DNA: A segment of DNA measuring 146 base pairs wraps approximately 1.65 to 1.67 times around this histone core in a left-handed direction, forming a structure known as a solenoidal supercoil.
- Linker DNA and Histone H1: Adjacent nucleosome cores are connected by a short segment of linker DNA. The fifth histone, H1, binds to this linker region, helping to stabilize the wrapping of DNA around the core and contributing to further compaction.
- Histone Tails: The amino-terminal tails of the core histones are intrinsically disordered and extend out from the nucleosome. These tails are primary sites for chemical modifications and are crucial for mediating interactions between adjacent nucleosomes, which facilitates their folding into more complex, higher-order structures.
- The formation of nucleosomes compacts the DNA length by approximately sevenfold, representing the first crucial step in chromosome packaging. This foundational unit serves as the building block for all subsequent levels of chromosome condensation.
3. From Simple Beads to Complex Chromosomes: Higher-Order Structure
While the sevenfold compaction achieved by nucleosome formation is significant, it is only the first step. To achieve the more than 10,000-fold compaction observed in a metaphase chromosome, chromatin must be folded through several additional hierarchical levels. This intricate folding must be highly organized to prevent the DNA from becoming an unmanageable, tangled knot.
- 3.1. Managing DNA Topology: The Role of Supercoiling and Topoisomerases The process of wrapping DNA around the histone core has important topological consequences. As illustrated in Figure 24-25, when a segment of DNA coils around the histone octamer, it introduces a negative supercoil into the bound DNA. Because the DNA is a closed loop (topologically), this change must be compensated by the formation of a positive supercoil in the adjacent, unbound DNA.
- This is where topoisomerase enzymes become essential. These enzymes act to relax the compensatory positive supercoils in the unbound DNA. By removing the positive supercoils, the topoisomerases leave a net negative supercoil, effectively trapping the DNA in an "underwound" state. This underwound state is energetically favorable for both further compaction and for processes that require strand separation, such as transcription and replication.
- 3.2. Advanced Folding and Nuclear Organization Beyond the level of individual nucleosomes, chromosomes are organized into even larger structures within the nucleus.
- Chromosomal Scaffold: The nucleosome fiber is further organized by associating with a proteinaceous chromosomal scaffold. This interaction arranges the DNA into a series of large loops.
- Compartments: The chromosome is partitioned into two major types of compartments. Transcriptionally active regions, known as euchromatin, have a more open, decondensed structure. In contrast, transcriptionally inactive regions, called heterochromatin, are highly condensed.
- Topologically Associating Domains (TADs): Within these compartments, the DNA is organized into large loops known as TADs, which average about 800,000 base pairs in length. These loops are often constrained at their base by the CCCTC-binding factor (CTCF) protein, which tethers distant DNA sites together.
- Chromosome Territories: Each individual chromosome is confined to a specific, discrete subnuclear domain known as a chromosome territory. There is very little intermingling of DNA between different territories, helping to maintain genomic organization.
The architectural organization of chromosomes is not fixed but is dynamically regulated throughout the life of the cell.
4. The Dynamic Nature of Chromatin
A key feature of chromosome structure is its dynamic nature. Far from being a static storage unit, chromatin must be accessible and adaptable to accommodate essential cellular processes. The degree of chromosome condensation changes dramatically throughout the cell cycle to facilitate DNA replication, gene expression, and the segregation of chromosomes during mitosis.
- 4.1. Chromosome Structure through the Eukaryotic Cell Cycle As shown in Figure 24-22, the state of chromatin varies significantly between the major phases of the cell cycle.
- Interphase: During this growth phase, chromatin is relatively decondensed and appears amorphous. In G1, the cell grows; in the S phase, the DNA is replicated to produce two identical sister chromatids; and in G2, the sister chromatids remain cohered.
- Mitosis: This is the phase of cell division, where chromatin undergoes its most dramatic condensation.
- Prophase: The replicated DNA undergoes extensive condensation, becoming visible as distinct chromosomes.
- Metaphase: The highly condensed chromosomes, each consisting of a pair of sister chromatids, align at the center of the cell.
- Anaphase: The sister chromatids separate and are pulled to opposite poles of the cell.
- Telophase: The separation process is completed, two new nuclei form, and the chromosomes begin to decondense, returning to their interphase state.
- 4.2. Orchestrating Change: SMC Proteins (Cohesins and Condensins) Alongside histones and topoisomerases, a third major class of chromatin-associated proteins, the SMC (structural maintenance of chromosomes) proteins, are critical for orchestrating these large-scale structural changes. The two major types of eukaryotic SMC proteins are cohesins and condensins.
|
Feature |
Cohesins (SMC1-SMC3) |
Condensins (SMC2-SMC4) |
|
Primary Role |
Function to link sister chromatids together immediately after DNA replication, holding them in close association. |
Drive the condensation of chromosomes as the cell prepares to enter mitosis, maintaining them in a compact state. |
|
Timing of Action |
Loaded onto chromosomes during the G1 phase. They are removed during anaphase by the enzyme separase to allow sister chromatid separation. |
Bind to chromosomes at the onset of mitosis to promote and maintain the highly condensed structure required for segregation. |
- 4.3. Epigenetics and Regulation: Histone Variants and lncRNAs Regulation of chromatin structure extends beyond the standard proteins to include heritable information that is not encoded in the DNA sequence itself. This epigenetic information includes the placement of histone variants and the action of regulatory RNA molecules. The covalent modifications mentioned earlier—such as acetylation and methylation—are the primary mechanisms that create these heritable epigenetic marks.
- Histone Variants: In addition to the standard histones, cells produce variant forms (e.g., H3.3, H2AZ, H2AX, CENPA) that can be incorporated into nucleosomes. These variants create specialized chromatin regions tailored for specific functions. For example, H3.3 and H2AZ are often found in regions of active transcription, H2AX is associated with DNA repair, and CENPA is associated with the repeated DNA sequences in centromeres.
- Long Noncoding RNAs (lncRNAs): These functional RNA molecules, which do not code for proteins, can act as structural scaffolds. A powerful example is Xist RNA in female mammals. This large lncRNA is expressed from one of the two X chromosomes and coats it entirely. This coating action recruits proteins that orchestrate the large-scale condensation and inactivation of that entire chromosome, a process that ensures proper gene dosage.
These biological mechanisms are not only fundamental to cell biology but also have direct relevance to medicine and provide fascinating points of comparison across different domains of life.
5. Clinical and Comparative Perspectives
Understanding the intricate architecture of chromosomes is not merely an academic exercise. This knowledge has profound implications for human health, particularly in the development of cancer therapies. Furthermore, comparing the strategies used by different organisms, such as bacteria and eukaryotes, to organize their genomes reveals fundamental principles of biological design.
- 5.1. Medical Relevance: Targeting Topoisomerases in Cancer Therapy The enzyme topoisomerase II is essential for managing the topological changes required for DNA replication and chromosome segregation. Because cancer cells are characterized by rapid and uncontrolled division, they have elevated levels of this enzyme, making it an excellent target for chemotherapy.
- Quinolones (e.g., Ciprofloxacin): These are antibiotics that are highly selective for bacterial topoisomerases, making them effective against bacterial infections without harming human cells.
- Camptothecin derivatives (e.g., Topotecan): These drugs inhibit human type I topoisomerases. Two important derivatives are irinotecan (Campto) and topotecan (Hycamtin)—used to treat colorectal cancer and ovarian cancer, respectively.
- Other agents (e.g., Doxorubicin, Etoposide): These compounds are potent inhibitors of human type II topoisomerases and are used to treat a variety of tumors. They act by stabilizing the complex where the enzyme has cleaved the DNA, preventing religation and leading to DNA damage and cell death.
- 5.2. A Different Strategy: The Bacterial Nucleoid Bacteria, which lack a nucleus, package their circular chromosome into a compact structure called the nucleoid. This organization differs significantly from the histone-based chromatin of eukaryotes.
- Organization: The bacterial chromosome is organized into approximately 500 looped domains that are topologically constrained. However, unlike in eukaryotes, the boundaries of these domains are not static and are thought to move along the DNA.
- Proteins: Bacteria lack the nucleosomes found in eukaryotes. Instead, they possess abundant "histone-like" proteins (such as HU) that bind to DNA. These proteins associate and dissociate rapidly, creating a much more dynamic chromosome structure.
- Functional Implications: This dynamic and more accessible structure is well-suited to the bacterial lifestyle. Bacteria have much shorter cell cycles and higher metabolic rates than most eukaryotic cells. Their chromosome organization reflects the need for ready and rapid access to genetic information for continuous transcription and replication.
In conclusion, the structure of a chromosome is a masterwork of molecular engineering. It demonstrates how a vast amount of information encoded in a linear DNA molecule can be elegantly organized at multiple hierarchical levels. This organization achieves a remarkable balance, allowing for extreme compaction when necessary for cell division while remaining sufficiently dynamic and accessible to permit the expression and replication of genetic information—the very processes that define life.
Resources
- MIND_MAP
- FLASHCARDS
- SELF-ASSESSMENT
- PRESENTATION_SLIDES
{kind=link}