15 RNA METABOLISM
15 RNA METABOLISM: A Structured Learning Summary
Executive Summary
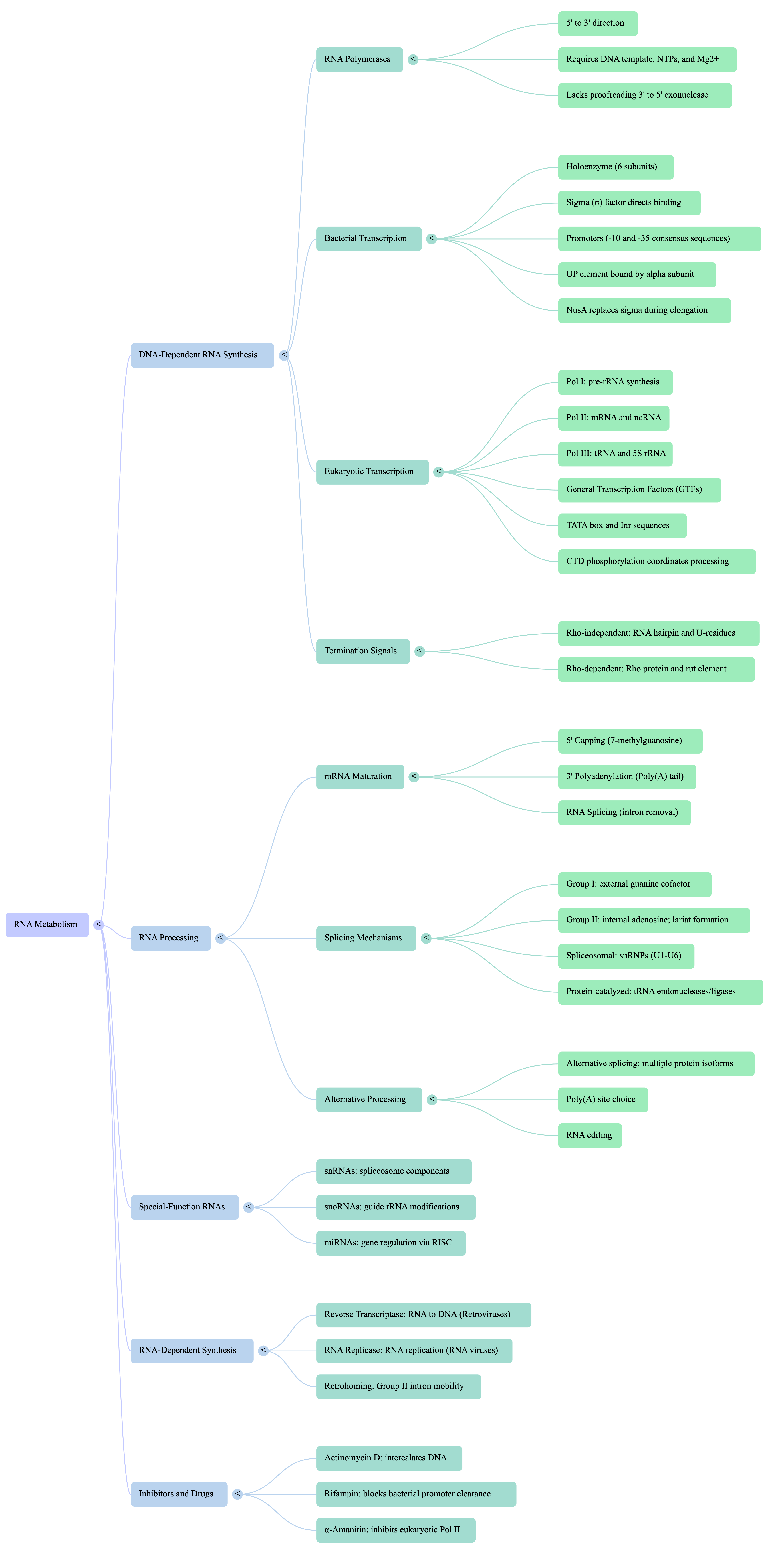
Ribonucleic acid (RNA) is a central molecule in the expression of genetic information, serving as far more than a simple messenger between DNA and protein. Its diverse roles as an information carrier, a biological catalyst, and a regulator of gene expression place it at the heart of cellular function. The primary process of RNA metabolism is transcription, the synthesis of an RNA molecule from a DNA template, catalyzed by the enzyme RNA polymerase. This process is highly selective, transcribing only specific segments of the genome into functional RNA molecules.
In eukaryotes, the initial RNA transcripts undergo a series of crucial modifications to become mature and functional. These post-transcriptional processing events include the addition of a protective 5' cap, the removal of non-coding sequences (splicing), and the attachment of a 3' poly(A) tail. These modifications are essential for the stability of messenger RNA (mRNA), its transport from the nucleus to the cytoplasm, and its efficient translation into protein.
The remarkable discovery that RNA itself can possess catalytic activity—acting as enzymes called ribozymes—revolutionized our understanding of molecular biology. This dual capacity to both store genetic information (like DNA) and catalyze chemical reactions (like proteins) provides the foundation for the "RNA World" hypothesis. This compelling theory posits that RNA, not DNA or protein, was the central macromolecule of early life, capable of storing a genetic blueprint while also catalyzing its own replication, thus solving the classic "chicken-and-egg" problem of which came first: the genetic blueprint or the functional machinery.
--------------------------------------------------------------------------------
1. The Fundamentals of RNA: More Than a Messenger
While DNA serves as the cell's permanent, stable archive of genetic information, RNA is the dynamic and versatile molecule responsible for actively expressing that information. All cellular RNA molecules are derived from information permanently stored in DNA, with the notable exception of the RNA genomes found in certain viruses. These transcripts are involved in a wide array of cellular processes, including acting as templates for protein synthesis and performing critical structural, catalytic, and regulatory functions.
1.1. The Four Major Classes of RNA
Cells produce four primary types of RNA, each with a distinct role:
- Messenger RNA (mRNA): These molecules encode the amino acid sequences of one or more polypeptides. They act as the genetic template that is read by the ribosome during protein synthesis.
- Transfer RNA (tRNA): tRNAs are adapter molecules that "read" the information encoded in the mRNA. For each three-nucleotide codon on the mRNA, the corresponding tRNA delivers the correct amino acid to the growing polypeptide chain.
- Ribosomal RNA (rRNA): These are the most abundant RNAs in the cell. They serve as structural and catalytic components of the ribosome, the complex molecular machine that carries out protein synthesis.
- Non-coding RNA (ncRNA): This is a broad and diverse category of RNA molecules that are not translated into protein. Instead, they perform a wide range of regulatory, structural, and catalytic functions. In the human genome, ncRNAs are the predominant products of transcription; while only about 2% of the genome codes for protein, approximately 76% is transcribed into RNA, much of it ncRNA.
1.2. The Transcriptome Concept
The complete set of RNA molecules produced in a cell under a given set of conditions is known as the transcriptome. Unlike the relatively static genome, the transcriptome is highly dynamic and provides a snapshot of which genes are actively being expressed at a particular moment. Analyzing the transcriptome reveals which segments of the genome are being utilized to meet the cell's immediate physiological needs.
These diverse RNA molecules are all synthesized through the fundamental process of transcription.
2. DNA-Dependent RNA Synthesis: The Process of Transcription
Transcription is the core process by which the static genetic information stored in a segment of DNA is converted into a functional RNA molecule. While it shares a fundamental chemical mechanism with DNA replication, transcription is distinct in several key ways. It does not require a primer to begin, it typically copies only limited segments of the DNA molecule, and for any given gene, only one of the two DNA strands serves as the template.
2.1. The Core Machinery: RNA Polymerase
The central enzyme of transcription is RNA polymerase. This enzyme moves along the DNA template, locally unwinding the double helix to form a "transcription bubble." As it proceeds, it synthesizes a complementary RNA strand. This movement generates positive supercoils (overwinding) in the DNA ahead of the enzyme and negative supercoils (underwinding) behind it.
2.2. Transcription in Prokaryotes (E. coli Model)
The process of transcription in the bacterium E. coli provides a foundational model for understanding this process. It occurs in three main stages: initiation, elongation, and termination.
- Initiation at Promoters: Transcription begins at specific DNA sequences called promoters. In E. coli, the most common class of promoters is recognized by an RNA polymerase holoenzyme containing the σ⁷⁰ subunit. These promoters contain two key consensus sequences:
- The -10 region (sequence: TATAAT)
- The -35 region (sequence: TTGACA)
- In the promoters of highly expressed genes, an additional UP (upstream promoter) element is often present, which further enhances binding of the RNA polymerase.
- Elongation: Once initiated, the RNA transcript is extended at a rate of 50 to 90 nucleotides per second.
- Termination: Transcription ceases at specific termination signals, of which there are two main classes in E. coli:
- ρ-independent termination: This mechanism depends on two sequence features. The first is a region that, when transcribed, forms a stable hairpin structure in the nascent RNA. The second is a downstream string of adenine (A) residues in the DNA template strand. The formation of the hairpin is thought to physically pull the RNA transcript away from the DNA template, and this tension, combined with the inherently weak U-A base pairing at the RNA-DNA hybrid, causes the transcript to dissociate from the polymerase.
- ρ-dependent termination: This process requires a protein factor called ρ (rho). The ρ protein binds to a specific CA-rich sequence on the nascent RNA transcript called a rut (rho utilization) element. It then migrates along the RNA toward the RNA polymerase, where it acts as a helicase to separate the RNA transcript from the DNA template.
2.3. Transcription in Eukaryotes: A More Complex System
Transcription in eukaryotes is significantly more complex, involving multiple RNA polymerases and a large suite of accessory proteins called transcription factors. The synthesis of mRNA is carried out by RNA Polymerase II (Pol II).
- Eukaryotic RNA Polymerase II (Pol II): The eukaryotic Pol II is a large, 12-subunit enzyme. Its largest subunit, RBP1, possesses a unique and crucial feature: a long carboxyl-terminal domain (CTD). This CTD consists of many repeats of a seven-amino-acid sequence (YSPTSPS) that acts as a regulatory hub.
- Initiation and General Transcription Factors: Unlike in bacteria, eukaryotic Pol II cannot bind directly to promoter DNA. It requires a set of proteins called general transcription factors to assemble at the promoter and form a preinitiation complex (PIC). This assembly occurs in a specific order:
- TFIID (containing TATA-binding protein, TBP) binds to the promoter.
- TFIIA and TFIIB bind sequentially.
- TFIIF, already associated with Pol II, is recruited to the complex.
- TFIIE and TFIIH bind to complete the assembly of the closed PIC.
- The Role of TFIIH: The TFIIH factor has two critical functions. Its helicase activity unwinds the DNA at the transcription start site to create the open complex, and its kinase activity phosphorylates the Pol II CTD. This phosphorylation triggers a conformational change that allows the polymerase to clear the promoter and begin elongation.
- Elongation and Termination: The phosphorylation state of the Pol II CTD changes dynamically, creating a binding code that recruits different factors at each stage of transcription. Phosphorylation at Ser5 of the heptad repeat marks the CTD for recruitment of capping enzymes during initiation. As the polymerase transitions to elongation, Ser2 is also phosphorylated, creating a new binding surface for splicing and polyadenylation factors. After termination, Pol II is dephosphorylated and can be recycled for another round of transcription.
Once this primary transcript is synthesized, particularly in eukaryotes, it is not yet ready for its final function and must undergo extensive processing.
3. Post-Transcriptional Processing: Maturing the RNA Transcript
The primary RNA transcripts synthesized during transcription, especially the pre-mRNAs in eukaryotes, are often non-functional and must be chemically modified to become mature RNAs. This series of modifications, known as RNA processing, ensures the stability of the RNA, facilitates its transport from the nucleus to the cytoplasm, and is essential for its correct biological function.
3.1. Eukaryotic mRNA Processing: A Three-Part Process
Nearly all eukaryotic mRNAs undergo three coordinated processing events before they are exported from the nucleus. These are not isolated events; they are physically and functionally coupled to transcription, as the enzymes responsible for each step are tethered to the carboxyl-terminal domain (CTD) of RNA Polymerase II, which acts as a moving platform coordinating the entire process.
- The 5' Cap:
- Structure: A 7-methylguanosine residue is attached to the 5' end of the mRNA via an unusual 5',5'-triphosphate linkage.
- Importance: The 5' cap serves two primary functions. First, it protects the mRNA from degradation by ribonucleases that attack the 5' end. Second, it is recognized by specific cap-binding proteins that are essential for the binding of the mRNA to the ribosome to initiate translation.
- Splicing: Removing Introns:
- Eukaryotic genes are often fragmented. They contain coding sequences called exons that are interrupted by non-coding intervening sequences called introns.
- Splicing is the process that removes introns and joins the exons together to form a continuous coding sequence. This reaction is catalyzed by a large and dynamic ribonucleoprotein complex called the spliceosome, which is composed of several small nuclear RNAs (snRNAs) and numerous proteins that together form small nuclear ribonucleoproteins (snRNPs).
- Significantly, some introns, known as Group I and Group II introns, can catalyze their own removal without the help of any protein enzymes. The discovery of these self-splicing introns was a landmark in biology, as it demonstrated that RNA could function as a biological catalyst—a ribozyme.
- The 3' Poly(A) Tail:
- Structure: A long chain of 80 to 250 adenine nucleotides, the poly(A) tail, is added to the 3' end of the mRNA.
- Process: This process is not templated by the DNA. The primary transcript is first cleaved downstream of a consensus signal sequence (AAUAAA). Then, the enzyme polyadenylate polymerase adds the long string of A residues.
- Function: The poly(A) tail, along with its associated binding proteins, helps protect the mRNA from enzymatic degradation in the cytoplasm and plays a role in coordinating transcription termination with the initiation of translation.
3.2. Processing of Other RNA Types (tRNA and rRNA)
Messenger RNAs are not the only transcripts that require processing.
- tRNAs and rRNAs are also synthesized as longer precursor transcripts that must be cleaved to their mature lengths. Processing often includes the chemical modification of bases, such as methylation or the conversion of uridine to pseudouridine. The 5' end of tRNA precursors is precisely cleaved by the ribozyme RNase P. For tRNAs, a conserved CCA sequence is added to the 3' end, which is the site of amino acid attachment.
- In eukaryotes, the modification of rRNA nucleosides is guided by another class of non-coding RNAs called small nucleolar RNAs (snoRNAs). These snoRNAs are part of large complexes (snoRNPs) that identify specific sites on the pre-rRNA for modification.
This intricate processing machinery can also be regulated to produce different outcomes from the same initial transcript, a concept known as alternative processing.
4. Generating Diversity: Alternative RNA Processing
One of the great paradoxes of modern genomics is that the complexity of an organism does not correlate with its number of genes. The biochemical solution to this paradox lies in differential RNA processing, a set of powerful mechanisms that allow a single gene to encode multiple distinct proteins. By generating variety from a fixed set of genetic instructions, these processes vastly expand an organism's functional capacity.
4.1. Alternative Splicing
Alternative splicing is a regulated process in which a particular exon may be either included in or excluded from the final mature mRNA. This generates multiple mRNA "isoforms" from a single pre-mRNA, which are then translated into different proteins with potentially different functions. This mechanism is remarkably prevalent, occurring in the transcripts of over 95% of human genes.
- Case Study: Spinal Muscular Atrophy (SMA)
- SMA is a severe neurodegenerative disease caused by a defect in the SMN1 gene, which prevents the production of the essential SMN protein.
- Humans have a nearly identical backup gene called SMN2. However, due to a subtle sequence difference, the SMN2 pre-mRNA is typically spliced in a way that excludes exon 7, resulting in an unstable and non-functional protein.
- The drug nusinersen is a revolutionary treatment for SMA. It is an antisense oligonucleotide (ASO) designed to bind to a specific silencer sequence on the SMN2 pre-mRNA. This binding masks the silencer from the splicing machinery, forcing the inclusion of exon 7. As a result, the SMN2 gene is able to produce a full-length, functional SMN protein, effectively compensating for the defective SMN1 gene and halting the progression of the disease.
4.2. Poly(A) Site Choice
Another mechanism for generating protein diversity is poly(A) site choice. If a primary transcript contains more than one polyadenylation signal site, the cell can choose where to cleave and polyadenylate the transcript. This produces mRNAs with different 3' ends, which can lead to proteins with different C-termini.
A classic example is the processing of the calcitonin gene transcript, which produces two different hormones in a tissue-specific manner:
|
Tissue |
Processing Outcome |
Final Product |
|
Thyroid |
Exon 4 is retained; cleavage at the first poly(A) site. |
Calcitonin |
|
Brain |
Exon 4 is spliced out; cleavage at the second poly(A) site. |
CGRP |
Together, alternative splicing and poly(A) site choice are powerful mechanisms that dramatically increase the protein diversity encoded by the genomes of higher eukaryotes.
5. Beyond the Central Dogma: RNA-Dependent Nucleic Acid Synthesis
The central dogma of molecular biology describes the flow of genetic information from DNA to RNA to protein. However, this model is an oversimplification. Certain enzymes can use RNA as a template to synthesize either DNA or additional RNA, revealing a more flexible and complex system of information transfer with profound implications for virology, chromosome biology, and our understanding of evolution.
5.1. Reverse Transcriptase: From RNA to DNA
Reverse transcriptase is an RNA-dependent DNA polymerase first discovered in retroviruses. These viruses, such as HIV, carry their genetic information in the form of RNA.
- Viral Life Cycle: Upon infecting a host cell, the reverse transcriptase uses the single-stranded viral RNA genome as a template to synthesize a complementary DNA (cDNA) strand. It then synthesizes a second DNA strand, creating a double-stranded DNA copy of the viral genome, which is subsequently integrated into the host cell's chromosome.
- Clinical Significance: Reverse transcriptases have a notably high error rate. The HIV enzyme is particularly error-prone—about 10 times more so than other known reverse transcriptases—introducing approximately one mutation per 20,000 nucleotides. This leads to the rapid mutation and evolution of HIV, which complicates the development of effective vaccines.
- Therapeutic Targeting: The anti-HIV drug AZT (azidothymidine) works by targeting this enzyme. AZT is an analog of thymidine that, when incorporated into the growing viral DNA chain, acts as a chain terminator. Because it lacks the 3'-hydroxyl group required to form the next phosphodiester bond, its incorporation permanently halts DNA chain extension. Since HIV reverse transcriptase has a higher affinity for AZT than cellular DNA polymerases do, the drug can selectively inhibit viral replication.
5.2. Telomerase: Solving the End-Replication Problem
Linear eukaryotic chromosomes face a challenge during replication: the very ends of the DNA cannot be fully copied by conventional DNA polymerases, which would lead to a progressive shortening of the chromosomes with each cell division.
- Function: Telomerase is a specialized reverse transcriptase that solves this "end-replication problem." This enzyme is a ribonucleoprotein that contains its own RNA template. It uses this template to add short, repetitive DNA sequences, known as telomeres, to the 3' ends of chromosomes.
- Cellular Aging: Telomerase is highly active in germ-line cells, ensuring that full-length chromosomes are passed on to the next generation. However, it is absent from most somatic (non-germ-line) cells. The gradual shortening of telomeres in these cells is linked to cellular senescence and the overall aging process.
These RNA-dependent processes suggest that RNA has played a central role not only in the modern cell but also in the evolutionary origins of life itself.
6. The RNA World Hypothesis: A Glimpse into the Origin of Life
The diverse functions of RNA—as an information carrier, a catalyst, and a regulator—provide compelling evidence for the "RNA World" hypothesis. This theory proposes a solution to the classic "chicken-and-egg" problem of early life: which came first, the genetic information (DNA) or the catalytic machinery (proteins)? The RNA World hypothesis suggests that the answer is neither. Instead, life may have begun with RNA, a single molecule capable of performing both roles.
The discovery of ribozymes—RNA molecules with catalytic activity, such as self-splicing introns and RNase P—was a crucial milestone supporting this theory. In a prebiotic world, an RNA molecule could have stored the blueprint for its own structure while also catalyzing its own replication, making it the most plausible candidate for the first self-replicating macromolecule. This dual functionality elegantly bridges the conceptual gap between simple organic chemistry and the complex, DNA- and protein-based cellular life we see today.
Resources
- MIND_MAP
- FLASHCARDS
- SELF-ASSESSMENT
- PRESENTATION_SLIDES
{kind=link}