09 REGULATORY ENZYMES
09 REGULATORY ENZYMES
1. Executive Summary
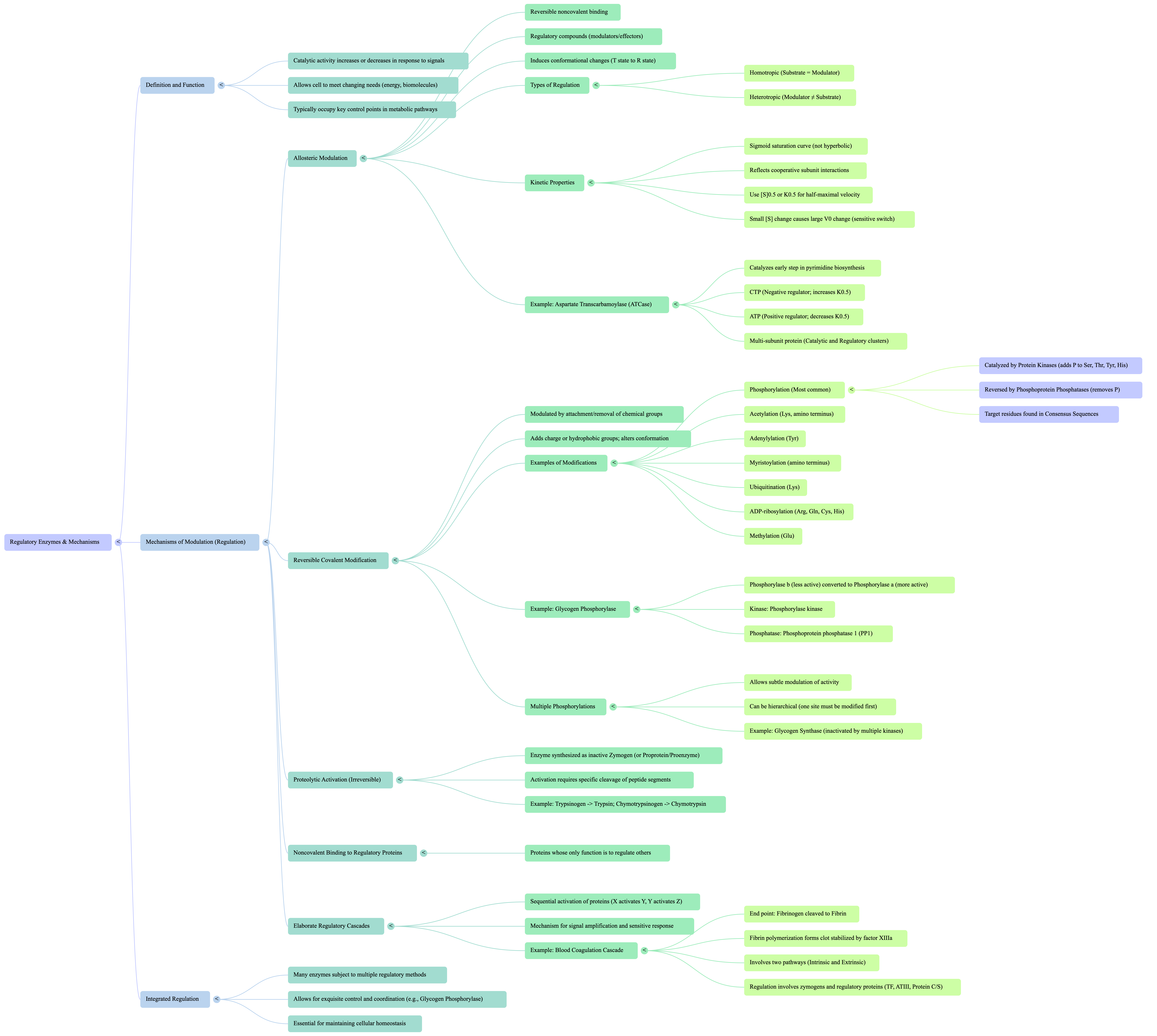
The persistence and adaptability of life depend on the precise control of countless biochemical reactions occurring within every cell. While enzymes are the catalysts that make these reactions possible, it is the regulation of their activity that orchestrates the complex symphony of metabolism. This document outlines the fundamental principles of enzyme regulation, explaining how cells dynamically adjust their internal chemistry to meet changing needs for energy, grow, and maintain a stable internal environment (homeostasis).
Cells employ four primary strategies to modulate enzyme activity. Allosteric regulation acts like a dimmer switch, where small molecules called modulators bind to a site on the enzyme separate from the active site, causing conformational changes that either increase or decrease its activity. Reversible covalent modification, most commonly the addition and removal of phosphoryl groups, functions as a rapid on/off switch, altering an enzyme's structure and function. Proteolytic cleavage offers an irreversible "on" switch, where inactive enzyme precursors, called zymogens, are permanently activated by the removal of a peptide segment; this is a common strategy for digestive enzymes and in the blood coagulation cascade. Finally, regulatory cascades create a powerful signal amplification system, where a small initial signal triggers a sequence of enzyme activations, leading to a massive and swift cellular response.
Crucially, these mechanisms are not mutually exclusive. Many key enzymes are subject to multiple layers of control, integrating signals from various metabolic pathways. This integration allows for exquisitely fine-tuned responses to complex cellular conditions. Understanding these regulatory mechanisms is fundamental to comprehending how cells manage resources, respond to their environment, and sustain the highly ordered processes we call life.
2. The Critical Role of Enzyme Regulation in Cellular Metabolism
Cellular metabolism is not a chaotic collection of independent reactions but a highly organized network of sequential pathways, where the product of one enzyme becomes the substrate for the next. Within these pathways, certain enzymes, known as regulatory enzymes, serve as critical control points. The catalytic activity of these enzymes can be increased or decreased in response to specific signals, thereby controlling the overall rate, or flux, of the entire metabolic sequence. This control is not merely a matter of efficiency; it is a fundamental requirement for survival. It allows the cell to economize its resources, respond to fluctuating demands for energy and biosynthetic building blocks, and prevent the wasteful synthesis or breakdown of essential molecules.
Enzyme regulation is vital for the cell for several key reasons:
- Meeting Changing Energy Needs: Cells must adjust the rate of catabolic (breakdown) pathways to generate ATP when energy is scarce and slow them down when energy is plentiful.
- Providing Biomolecules for Growth and Repair: The synthesis of amino acids, nucleotides, and lipids must be coordinated with the cell's growth and maintenance requirements.
- Ensuring Pathway Efficiency: Regulation ensures that groups of enzymes in a sequential pathway work together harmoniously, avoiding bottlenecks or the accumulation of unnecessary intermediates.
- To prevent chaos by ensuring that specific, complex macromolecules are not wastefully broken down, thereby maintaining cellular structure and order. This control creates the "complex, highly regulated symphony we call life."
To achieve this sophisticated level of control, cells have evolved several distinct and powerful mechanisms for modulating enzyme activity.
3. Key Mechanisms of Enzyme Regulation
Cells utilize several distinct strategies to finely tune the activity of their enzymes. Each mechanism offers unique advantages in terms of speed, reversibility, and the type of signals it can respond to. The four major regulatory strategies that orchestrate cellular metabolism are allosteric regulation, reversible covalent modification, proteolytic activation, and regulatory cascades.
3.1 Allosteric Regulation: The Body's Dimmer Switch
Allosteric regulation involves the reversible, noncovalent binding of a regulatory molecule—called an allosteric modulator or effector—to a site on the enzyme other than the active site. This secondary location is known as the allosteric site. The binding of a modulator induces a conformational change in the enzyme, which in turn alters the shape and catalytic efficiency of the active site.
- Conformational States: Allosteric enzymes can interconvert between two conformations: a less-active form, often called the T state (for tense), and a more-active form, the R state (for relaxed). The balance between these two states determines the enzyme's overall activity.
- Modulators: A positive modulator (activator) is a molecule that binds to and stabilizes the active R state, increasing the enzyme's activity. Conversely, a negative modulator (inhibitor) binds to and stabilizes the inactive T state, reducing enzyme activity.
- Homotropic vs. Heterotropic Regulation: This mechanism is further classified by the identity of the modulator. In homotropic regulation, the substrate itself acts as a positive modulator by binding to the active site, which also serves as the regulatory site. This binding increases the catalytic activity of the enzyme's other active sites. In heterotropic regulation, the modulator is a molecule different from the substrate.
Kinetic Properties of Allosteric Enzymes
Unlike enzymes that follow simple Michaelis-Menten kinetics, allosteric enzymes exhibit distinct kinetic behavior.
- A plot of initial reaction velocity (V₀) versus substrate concentration ([S]) for an allosteric enzyme typically produces a sigmoid (S-shaped) saturation curve, rather than a hyperbola.
- Because the kinetics are not hyperbolic, the term Km is not used. Instead, K₀.₅ (or [S]₀.₅) denotes the substrate concentration that yields half-maximal velocity.
- The sigmoid curve provides a significant biological advantage. In the steep part of the curve, a small increase in substrate concentration can cause a large increase in the reaction velocity, creating a highly sensitive molecular switch.
The binding of heterotropic modulators alters the shape of this curve, allowing for fine-tuned control.
|
Modulator Type |
Effect on K₀.₅ |
Effect on Vmax |
Curve Shape |
|
Positive (Activator) |
Decreases K₀.₅ |
Typically unchanged |
Becomes more hyperbolic |
|
Negative (Inhibitor) |
Increases K₀.₅ |
Typically unchanged |
Becomes more sigmoidal |
|
Less Common Type |
Nearly constant |
Can be increased or decreased |
Vmax changes |
A classic example is Aspartate Transcarbamoylase (ATCase), which catalyzes an early step in pyrimidine biosynthesis. The pathway's end product, CTP, acts as a negative modulator, inhibiting ATCase to prevent overproduction. In contrast, ATP, which signals an abundance of energy and resources for growth, acts as a positive modulator, stimulating the enzyme's activity.
3.2 Reversible Covalent Modification: Chemical On/Off Switches
This regulatory strategy involves the attachment and removal of a chemical group to one or more amino acid residues on an enzyme, mediated by other specific enzymes. This covalent modification alters the enzyme's properties and, consequently, its activity.
- Common modifying groups include phosphoryl, acetyl, adenylyl, uridylyl, methyl, and ubiquitin. The attachment of these groups can introduce new properties, such as charge or hydrophobicity, that trigger conformational changes in the enzyme.
The most common and important type of covalent modification is phosphorylation.
- Protein kinases are enzymes that catalyze the transfer of a phosphoryl group from ATP to the hydroxyl group of a Serine (Ser), Threonine (Thr), or Tyrosine (Tyr) residue on a target protein.
- Phosphoprotein phosphatases reverse this process by catalyzing the removal of these phosphoryl groups.
- The addition of a bulky, negatively charged phosphoryl group can dramatically alter a protein's conformation and catalytic activity. Its negative charges can repel neighboring negatively charged residues (like Asp or Glu) or form new electrostatic interactions and hydrogen bonds, causing dramatic conformational changes.
A key example is Glycogen Phosphorylase, the enzyme responsible for breaking down glycogen into glucose 1-phosphate.
- It exists in a less active form, phosphorylase b, and a more active form, phosphorylase a.
- Phosphorylase kinase converts the b form to the a form by phosphorylating a specific Ser residue on each subunit.
- Phosphoprotein phosphatase 1 (PP1) reverses this activation by removing the phosphoryl groups.
Some enzymes, like Glycogen Synthase, have numerous phosphorylation sites. This multiple phosphorylation allows for extremely subtle and fine-tuned modulation of activity, where different combinations of phosphorylated sites produce a wide spectrum of activity levels, sometimes in a hierarchical fashion.
3.3 Proteolytic Activation: An Irreversible Commitment
Proteolytic cleavage is an irreversible mechanism used to activate certain enzymes. The enzyme is initially synthesized as an inactive precursor, known as a zymogen (or more generally, a proprotein or proenzyme).
The biological purpose of this strategy is to ensure that powerful enzymes, especially those with high destructive potential, are only activated in the correct location or at the appropriate time. For instance, digestive proteases like trypsin and chymotrypsin are synthesized as inactive zymogens (trypsinogen and chymotrypsinogen) in the pancreas and are only activated after they have been safely transported to the intestine. This prevents them from digesting the cells in which they were made. Similarly, processes that must be strictly localized, like blood clotting, rely on this mechanism.
Once a zymogen has been activated by proteolytic cleavage, the process cannot be reversed. The enzyme's activity can only be terminated by a separate inhibitor protein, such as the pancreatic trypsin inhibitor, that binds tightly to its active site.
3.4 Regulatory Cascades: Amplifying the Signal
A regulatory cascade is a sequence of reactions in which an enzyme activates another enzyme, which in turn activates a third, and so on.
The primary advantage of a cascade is signal amplification. Because each activated enzyme is a catalyst, it can act on many molecules of the next enzyme in the sequence. This creates an exponential increase in the signal at each step, allowing a very small initial stimulus to generate a large and rapid physiological response.
The blood coagulation cascade is a quintessential example of this mechanism.
- The ultimate goal is the conversion of the soluble plasma protein fibrinogen into insoluble fibrin fibers, which form the meshwork of a blood clot. This final step is catalyzed by the protease thrombin.
- Two interconnected pathways lead to fibrin formation. The extrinsic pathway is so named because it is initiated by tissue factor (TF), a protein that is "extrinsic" to the blood, becoming exposed only after vascular injury. The intrinsic pathway uses only components found "intrinsic" to the blood plasma.
- Both pathways consist of a series of zymogen activations. They converge on the activation of factor X, which then cleaves and activates prothrombin to form thrombin. This sequence perfectly illustrates the cascade principle, amplifying the initial signal of vascular injury into a robust clot.
Given the potential danger of uncontrolled clotting (thrombosis), the cascade is tightly regulated. Inhibitory proteins like antithrombin III (ATIII), which directly inactivates key proteases like thrombin, and protein C, which inactivates crucial regulatory cofactors, are vital for limiting the clot to the site of injury and shutting down the cascade once damage is repaired.
4. Synergy in Regulation: The Integrated Control of Metabolism
The regulatory mechanisms described are not isolated strategies; they often work in concert to provide sophisticated, multilayered control over an enzyme's activity. Many of the most critical enzymes in metabolism are regulated by a combination of these methods. For example, glycogen phosphorylase is controlled by both reversible covalent modification (it is activated by phosphorylation) and allosteric regulation (its activity is further boosted by AMP, a signal of low cellular energy).
This complexity provides an enormous biological advantage. By integrating multiple signals—allosteric modulators reflecting the cell's immediate metabolic state and covalent modifications responding to hormonal or long-term signals—the cell can make nuanced "decisions." This allows for the exquisite fine-tuning of metabolic pathways, ensuring that cellular resources are used with maximum efficiency and that the cell can adapt seamlessly to a constantly changing internal and external environment.
Ultimately, the control of catalysis is as critical to life as catalysis itself. It is through these intricate regulatory networks that the thousands of simultaneous reactions within a cell are coordinated into the complex, highly regulated symphony we call life.
Resources
- MIND_MAP
- FLASHCARDS
- SELF-ASSESSMENT
- PRESENTATION_SLIDES
{kind=link}